野百合堿對人肝竇內皮細胞的毒性機制研究
2020-06-11 07:49:22王蔚倩葉鉉玲熊愛珍王崢濤
中國藥理學通報 2020年6期
關鍵詞:檢測
王蔚倩,葉鉉玲,陳 巖,熊愛珍,楊 莉,王崢濤
(上海中醫藥大學 1.中藥研究所中藥標準化教育部重點實驗室暨上海市復方中藥重點實驗室、2.交叉科學研究院,上海 201203)
近年來,中藥的“毒性”問題已引起國際社會的關注和擔憂,繼“馬兜鈴酸”風波之后,另一大類毒性更強、波及面更廣的天然植物性毒素-吡咯里西啶生物堿(pyrrolizidine alkaloids, PAs),成為國際上關注的新熱點[1]。PAs廣泛分布于6000多種高等植物中,目前已經發現660余種PAs及其氮氧化物,其中近一半都被報道對人或者牲畜有毒性[2]。被PAs污染的谷物、牛奶、蜂蜜等食物及含PAs的傳統草藥、補劑、茶等均可對人類造成極大的危害。PAs可導致肝竇阻塞綜合征(hepatic sinusoidal obstruction syndrome,HSOS)、肝巨大細胞癥、肝纖維化壞死等嚴重的肝臟損傷,目前在臨床上尚無有效的預防和治療藥物。
HSOS是肝竇內皮細胞(sinusoidal endothelial cells,SECs)損傷致肝竇流出道阻塞所引起的肝內竇性門脈高壓,又稱肝小靜脈閉塞病(hepatic veno-occlusive disease,HVOD),臨床主要癥狀為肝腫大、肝區疼痛、腹水、黃疸、高血膽紅素等[3]。其發病機制可能與SEC內谷胱甘肽耗竭,一氧化氮消耗,基質金屬蛋白酶(matrix metalloproteinases,MMPs)和血管內皮生長因子(vascular endothelial growth factor,VEGF)的肝內表達增加,以及凝血因子的激活等有關[4]。HSOS缺乏特效治療,重癥患者常因多臟器功能衰竭而死亡,在發病早期及時識別以及早期干預至關重要[5]。SEC鋪襯在肝血竇內,表面富含窗孔,為肝竇和竇周間隙(Disse間隙)之間溶質的交換開放通道,可以通過改變窗孔直徑和數量調節肝血竇內外的物質交換與信息交流,參與形成血液—肝細胞屏障[6]。由于SEC是HSOS的最直接損傷部位,所以近年來相關機制研究開始關注肝竇內皮的損傷,并以此作為研究切入點。
野百合堿(monocrotaline,MCT)主要存在于豬屎豆屬植物中,是目前研究最多的毒性PAs之一,常用來在動物上建立HSOS模型[7]。MCT進入體內后會被肝臟中的細胞色素P450酶代謝活化為脫氫野百合堿(dehydromonocrotaline,DMCT),并迅速與細胞大分子如蛋白質作用形成吡咯蛋白加合物(pyrrole-protein adducts,PPA),繼而引發肝毒性。有研究表明,DMCT與內皮細胞上的F-actin共價結合后,可導致肌動蛋白細胞骨架的解體和內皮細胞的圓形化甚至脫落,并伴隨著血紅細胞進入竇周間隙,同時導致MMPs的升高,從而降解細胞外基質,由此進一步造成SEC脫落[8-9]。SEC脫落死亡是HSOS發病過程中至關重要的因素,也是HSOS區別于其他類型肝損傷的顯著特征。
目前關于PAs對SEC毒性的報道研究大都利用整體動物模型,影響因素較多,而幾乎沒有將PAs直接作用于人肝竇內皮細胞(human hepatic sinusoidal endothelial cells,HHSECs)上進行體外毒性機制的深入研究。本研究以MCT為代表性藥物,研究其體外直接作用于HHSEC的毒性反應及其作用機制,為臨床HSOS的治療提供實驗基礎和理論依據。
1 材料
1.1 細胞與試劑HHSEC細胞株與其特制培養體系(MED-0002)購于中國武漢原生原代生物醫藥科技有限公司;人正常肝細胞株(L-02)購自中國科學院細胞庫;RPMI 1640培養基(批號2122752)購于美國Life Technology 公司;胎牛血清(批號1739463)及青霉素-鏈霉素(批號2068824)購于美國Gibco公司;MCT(BP0957)購自成都普瑞法科技開發有限公司; RIPA裂解液(批號TL277222)和BCA 蛋白測定試劑盒(批號UB276926)購于美國Thermo Fisher公司;抗體cleaved caspase-3(#9661)、PARP(#9532)、β-actin(#4970)購于美國Cell Signaling公司;還原型谷胱甘肽(GSH)試劑盒(批號20180821)購于南京建成生物科技有限公司;逆轉錄試劑盒(批號AI70509A)和實時定量PCR試劑盒(批號AJ10745A)購于日本TaKaRa公司;CCK-8試劑盒(批號PG678)購于日本同仁公司;Caspase-3熒光底物Ac-DEVD-AFC(批號2857919)購于美國Biomol公司;Hoechst染色試劑盒(批號022519190919)購于上海碧云天生物技術有限公司;RNA極速抽提試劑盒(#220011)購于上海飛捷生物技術有限公司。
1.2 儀器細胞培養箱(新加坡ESCO公司);SparkTM多功能酶標儀(瑞士TECAN公司);ABSCIEX QTRAP6500液相色譜-質譜聯用儀(美國ABSCIEX公司);凝膠電泳儀、電轉儀(美國Bio-Rad 公司);凝膠成像儀(美國GE 公司);熒光顯微鏡(日本OLYMPUS公司);QuantStudio 6 Flex 型實時熒光定量PCR儀(美國Applied Biosystems 公司)。
2 方法
2.1 藥物配制稱取適量MCT,用10%的鹽酸水溶液充分渦旋溶解,加1 mmol·L-1NaOH 調節至pH 7.2,配制母液500 mmol·L-1。
2.2 細胞培養將HHSEC培養于含5%滅活胎牛血清、1%內皮細胞生長添加劑和1%青霉素-鏈霉素的人肝竇內皮細胞特制培養基。人正常肝L-02細胞培養于含10%滅活胎牛血清和1%青霉素-鏈霉素的RPMI1640培養基中。細胞放置在37 ℃,5% CO2的培養箱中培養。
2.3 細胞活力檢測CCK-8試劑盒被用于檢測細胞活性。將HHSEC接種于96孔板,細胞貼壁后,將細胞分別與不同濃度的MCT,共孵育24 h后棄去原培養基,向每個孔中分別加入100 μL含10% CCK-8培養基,并在培養箱中孵育1 h,用酶標儀測定吸光度(450 nm)。將細胞存活率以對照組的百分比進行標準化并按以下公式計算細胞活力:細胞活力/%=(給藥組-空白組)/(對照組-空白組)×100%。
2.4 PPA含量測定參考文獻報道的方法,采用柱前衍生化LC-MS法檢測細胞及培養基中PPA的含量[10]。采用RIPA裂解液提取細胞總蛋白,細胞蛋白裂解液及培養基分別使用Ehrlich試劑反應衍生化處理,再利用液質連用儀進行檢測。以Waters Acquity BEH C18(2.1×100 mm,1.7 μm)色譜柱分離,0.1%甲酸溶液(A)-乙腈(B)為流動相梯度洗脫,梯度洗脫如下:0~1 min,10% B;1~6 min,40% B;6~10 min,95% B。流速為0.4 mL·min-1,進樣2 μL。采用多反應監測MRM(341>252;341>296)測定樣本中PPA的含量。
2.5 細胞內GSH含量測定將細胞與不同濃度的MCT共孵育24 h,收集細胞,參照GSH試劑盒說明書測定GSH含量。
2.6 Hoechst 33258細胞染色觀察將HHSEC接種于96孔板(2×104個/孔),細胞貼壁后分別加入不同濃度的MCT,共孵育24 h后棄去培養基,按照試劑盒說明書,加入固定液固定10 min,PBS洗滌兩遍后加入Hoechst 33258染色液染色5 min,再用PBS洗滌兩遍,在熒光顯微鏡下觀察。
2.7 Caspase-3酶活性檢測使用caspase-3熒光底物(Ac-DEVD-AFC)測定caspase-3活性。將細胞與不同濃度的MCT共孵育24 h,收集細胞,提取蛋白,用BCA 蛋白測定試劑盒測定蛋白濃度。參照試劑盒說明書,取20 μg蛋白,加20 μmol·L-1熒光底物及緩沖液至200 μL于96孔板中,混勻后在37 ℃恒溫箱中避光孵育1 h,測定熒光信號。熒光信號通過熒光酶標儀分別在400 nm和510 nm的激發波長和發射波長下檢測。根據給藥組與空白對照組吸光值的比值計算相對caspase-3活性。
2.8 蛋白免疫印跡法檢測將細胞與不同濃度的MCT共孵育24 h,收集細胞,提取蛋白,用BCA 蛋白測定試劑盒測定蛋白濃度。取適量蛋白變性,通過SDS-PAGE凝膠電泳分離蛋白(上樣量為每孔30 μg蛋白),然后電泳轉移至PVDF膜。先用5% BSA室溫封閉2 h,再與一抗(1 ∶ 1 000)4 ℃共孵育過夜,用PBST洗去殘留的一抗后與相應二抗(1 ∶ 5 000)孵育1 h,再用PBST洗去多余抗體,最后將膜放于顯影液中反應,置凝膠成像儀中觀察。以內參蛋白 β-actin為參照,計算目的蛋白的相對表達量。
2.9 實時熒光定量聚合酶鏈式反應分析將細胞與不同濃度的MCT共孵育24 h,收集細胞,根據RNAfast200總RNA極速抽提試劑盒說明書提取總RNA,用PrimeScript RT Master Mix試劑盒將1 μg RNA逆轉錄為cDNA。根據SYBR Premix Ex Taq試劑盒說明書使用SYBR green預混物進行實時熒光PCR檢測細胞中目的基因mRNA 表達水平。以內參基因GAPDH為參照,通過2-ΔΔCt方法分析目標基因的相對表達量。使用的引物序列見Tab 1。
Tab 1 Primers for qPCR analysis

3 結果
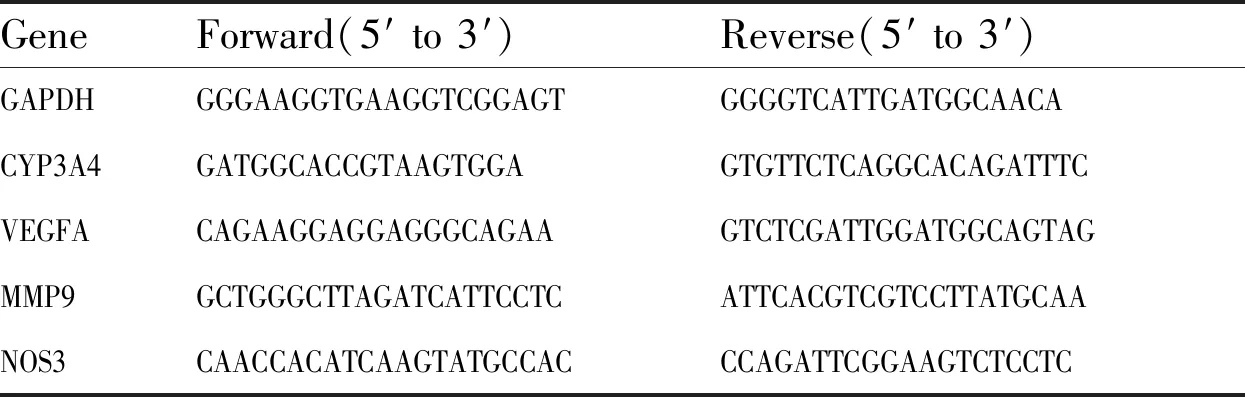
3.1 MCT對HHSEC細胞存活率的影響HHSEC與不同濃度MCT(0、0.5、1、5、10 mmol·L-1)共孵育24 h,通過CCK-8試劑盒檢測MCT對HHSEC細胞存活率的影響。由Fig 1A可知,與空白組比較,5 mmol·L-1MCT能顯著降低HHSEC的細胞存活率至72%,10 mmol·L-1MCT則將細胞存活率明顯降低為45%。然而,在測試濃度范圍內,MCT(0~10 mmol·L-1)對L-02細胞存活率無顯著影響(Fig 1B)。由此可知,HHSEC對MCT的毒性更加敏感。
3.2 細胞及培養基中PPA的含量測定HHSEC與不同濃度MCT(0、1、5 mmol·L-1)共孵育24 h后,檢測細胞及培養基中PPA的含量。由Fig 2可知,給藥后HHSEC細胞及培養基中都檢測到一定含量的PPA,且細胞及培養基中PPA含量均隨MCT給藥濃度增加而增多。
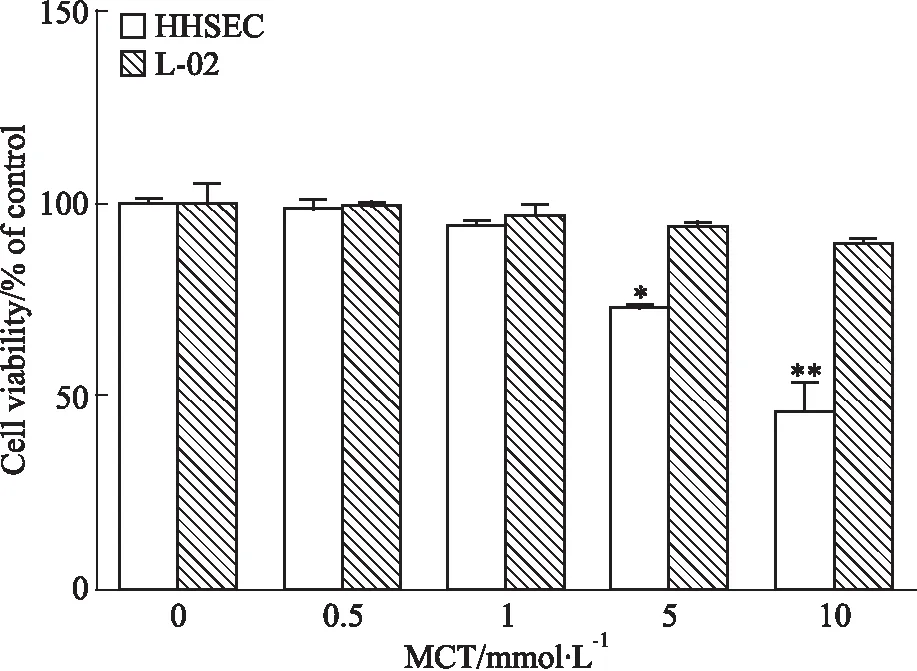
Fig 1 Cytotoxicity of MCT on HHSECs and L-02
*P<0.05,**P<0.01vs0 mmol·L-1MCT
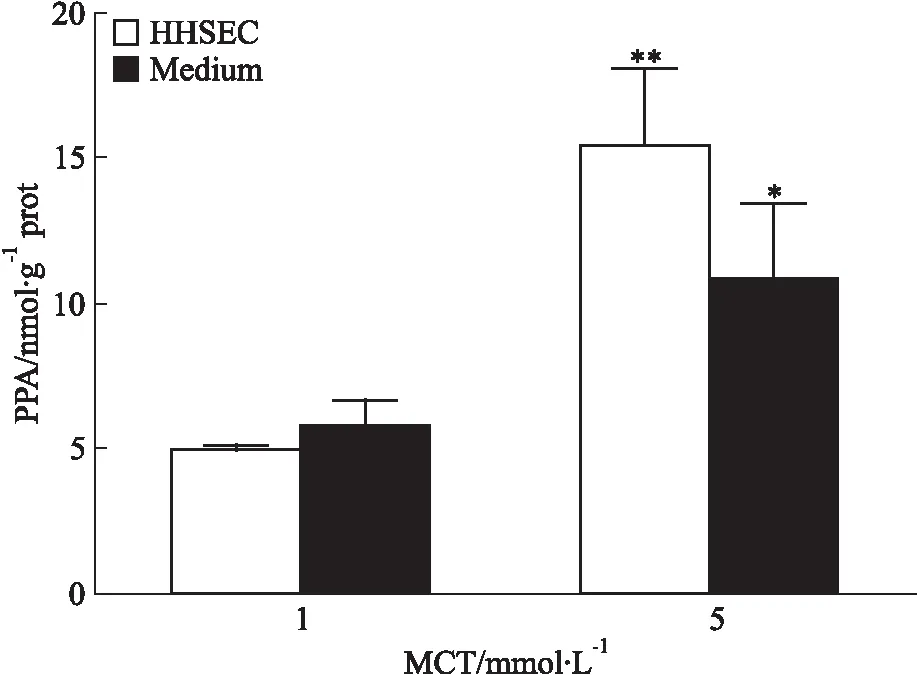
Fig 2 Concentration of PPA in cell lysate and medium of HHSECs exposed to
*P<0.05,**P<0.01vs1 mmol·L-1MCT.
3.3 MCT降低了HHSEC內GSH的含量HHSEC與不同濃度MCT(0、1、5 mmol·L-1)共孵育24 h后,HHSEC細胞內的GSH含量隨MCT給藥濃度的增加而減少。如Fig 3所示,5 mmol·L-1MCT給藥24 h可顯著降低細胞內GSH含量(P<0.05)。
3.4 MCT誘導HHSEC發生凋亡HHSEC與不同濃度MCT(0、5、10 mmol·L-1)共孵育24 h后,通過Hochest染色,caspase-3活性,cleaved caspase-3及cleaved parp的蛋白表達水平檢測細胞凋亡。由Hoechst33258染色結果可知(Fig 4A),5和10 mmol·
Fig 3 GSH content in HHSECs decreased by
*P<0.05vs0 mmol·L-1MCT
L-1MCT給藥組可見明顯細胞核固縮, 提示MCT可誘導HHSEC發生凋亡。5和10 mmol·L-1MCT給藥組caspase-3蛋白酶活性顯著升高,分別為空白對照組的3.0倍和17.8(Fig 4B)。由Fig 4C可知, MCT(10 mmol·L-1)給藥還可顯著增加HHSEC細胞凋亡活化蛋白標志物cleaved caspase-3及cleaved parp的蛋白表達水平。以上結果均表明,MCT可誘導HHSEC凋亡。
3.5 相關基因的表達水平檢測HHSEC與不同濃度MCT(0、1、5 mmol·L-1)共孵育24 h后,收集細胞并檢測了相關基因的mRNA表達水平。結果可知(Fig 5),相比空白對照組,5 mmol·L-1MCT給藥組CYP3A4基因表達水平升高3.9倍,VEGFA和MMP-9表達水平分別增加1.9倍和3.1倍,而NOS3表達水平則顯著下降,約為空白對照組的54%。
4 討論
PAs是導致臨床HSOS的主要原因之一[3]。近年來,因誤服誤用含PAs的中草藥及復方所致HSOS的報道逐年增多,引起了國際國內社會的廣泛關注。目前,關于HSOS的發病機制并不十分明確,涉及多種細胞、分子以及信號通路。大量研究表明,SEC損傷是HSOS發生的起始因素和關鍵環節[4,15],然而由于SEC細胞上代謝酶含量相對較低,幾乎沒有將PAs直接作用于SEC的體外研究,大多數研究者采用體內動物模型,使得其機制更加復雜。因此,本研究嘗試以HHSEC模型研究MCT的體外毒性作用,并初步探討其產生細胞毒性的機制。
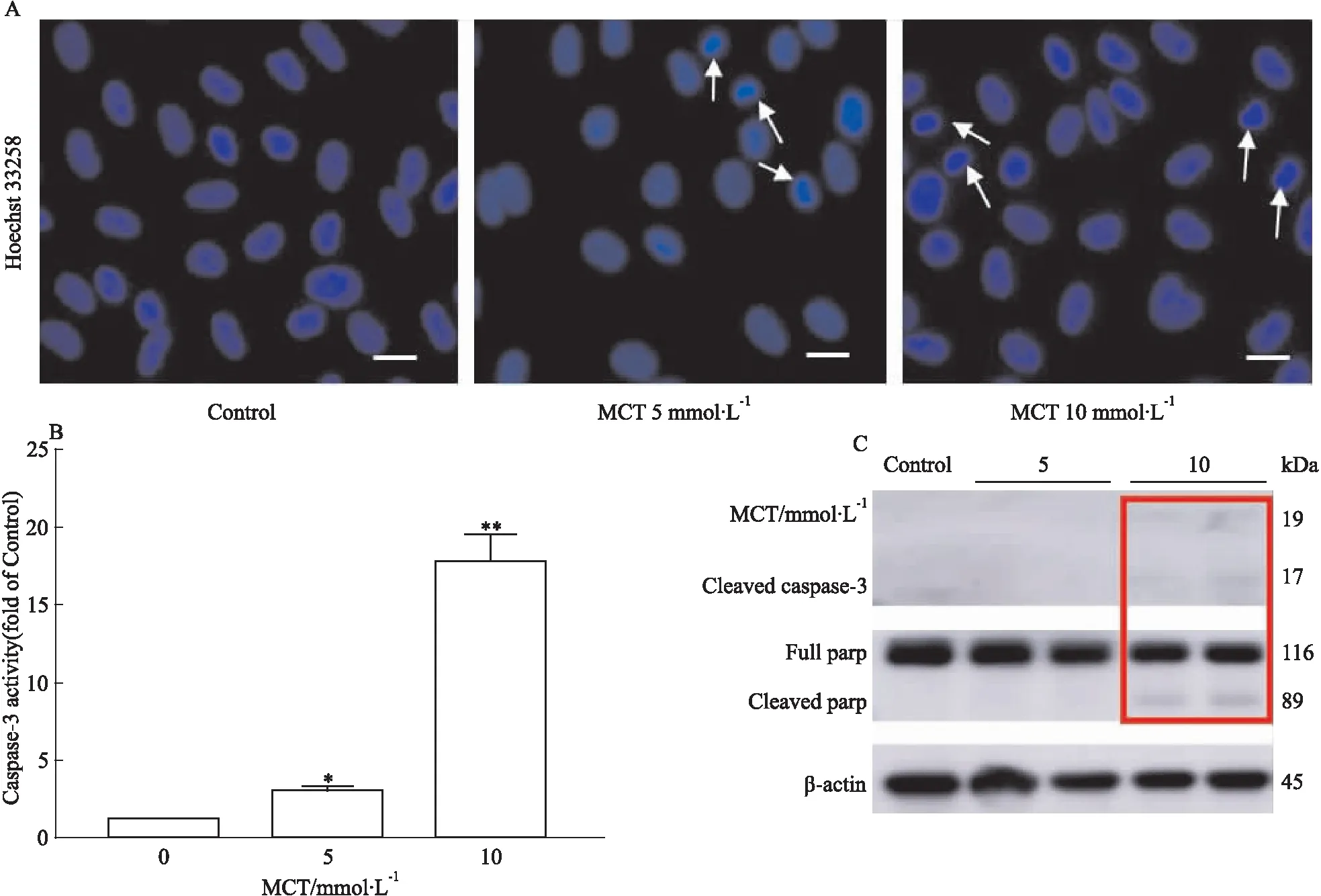
Fig 4 Apoptosis in HHSECs induced by
A:Representative images of Hoechst staining of HHSECs. White arrows denote apoptotic nuclei. The bar represents 25 μm;B:Caspase-3 activity assay of total cell lysates of HHSEC;C:The protein expression levels of cleaved caspase-3 and cleaved parp.*P<0.05,**P<0.01vs0 mmol·L-1MCT
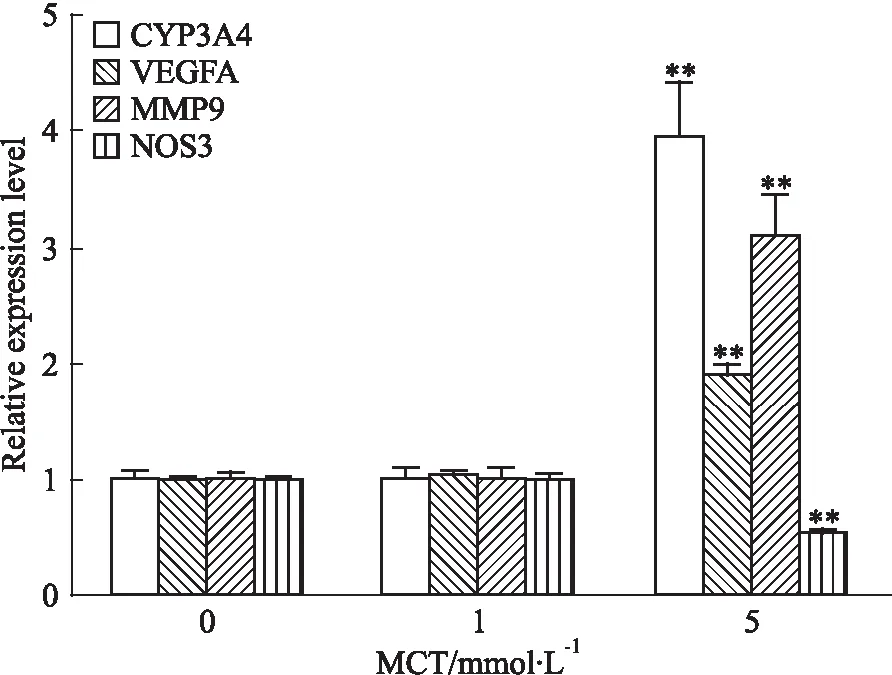
Fig 5 The mRNA expression levels of target genes in
**P<0.01vs0 mmol·L-1MCT
我們首先測試了MCT對HHSEC 的細胞毒性。結果表明MCT(5, 10 mmol·L-1)可顯著降低HHSEC的細胞存活率,而同樣濃度下MCT對L-02細胞則沒有明顯毒性,由此證明了MCT對HHSEC毒性的特異性。大部分PAs需過肝臟細胞色素P450酶(主要為CYP3A4酶)代謝活化并生成PPA繼而引發肝臟毒性。PPA不僅是PAs代謝活化的直接證據,也被中華醫學會消化病學分會肝膽疾病協作組建議作為PAs毒性生物標記物[3]。我們在給藥MCT的HHSEC細胞裂解液及培養基中均檢測到了大量PPA,且其含量與MCT給藥濃度且呈劑量依賴性增加。由此可知,MCT經HHSEC中的藥物代謝酶代謝活化后形成活性中間體,一方面與細胞中的蛋白結合形成PPA,另一方面也可排出細胞與培養基中的蛋白結合形成PPA。這與文獻在PAs所致HSOS的臨床患者及灌胃PAs的動物血液及肝組織檢測到PPA的報道相符[10]。我們進一步驗證了HHSEC細胞代謝酶的水平,在HHSEC中檢測到了一定含量的代謝活化關鍵酶CYP3A4,而MCT可以誘導CYP3A4的表達,進一步加速MCT代謝活化,從而加劇對HHSEC的損傷。
谷胱甘肽作為細胞內主要的抗氧化劑和自由基清除劑,對于細胞生理功能的維持具有重要作用。研究表明,PAs經代謝活化產生的活性中間體,除了與蛋白結合生成PPA進一步誘發毒性外,還可通過與GSH結合生成GSH結合物從而利于其排出而解毒[11],這也是體內PAs毒性的一條重要的減毒途徑。而HHSEC由于自身所含GSH水平較低,GSH大量消耗且無法及時補充,將使其更加易于受到毒物的侵害而死亡,從而加劇了HSOS損害[8]。與本實驗中,MCT給藥后HHSEC中GSH含量顯著下降,與文獻報道相符,也是造成其毒性的重要原因之一。
此外,細胞凋亡也被認為是PAs引發毒性的重要機制之一[12-14]。caspases蛋白家族在細胞凋亡過程中具有重要的調節作用。caspase-3蛋白是凋亡的關鍵執行者,在藥物或其他信號分子作用下,caspase-3被剪切活化產生cleaved caspase-3,并剪切其下游靶蛋白PARP產生cleaved parp,啟動細胞凋亡。因此,cleaved caspase-3及cleaved parp可用作凋亡的指標物。本實驗中,MCT給藥可誘導HHSEC細胞核固縮、深染,誘導caspase-3酶活力呈劑量依賴性增加,并誘導細胞中cleaved caspase-3及cleaved parp的蛋白表達水平升高。由此可知,MCT可誘導HHSEC發生凋亡,導致毒性。根據課題組前期的研究及文獻報道,PAs可在體內外誘導細胞凋亡導致毒性,但其中涉及到多種信號分子及作用機制,包括降解抗凋亡因子Bcl-xl[12]、誘導Drp1干擾線粒體分裂融合[13]、誘導Fas受體[14]等。然而,MCT誘導HHSEC凋亡具體通過哪些信號通路仍需進一步深入研究。
另有研究表明,MCT可引起大鼠竇狀隙內皮細胞中F-actin的解聚,導致SEC中金屬蛋白酶-9(MMP9)表達增加,從而降解細胞外基質造成SEC脫落誘導HSOS[15]。而Iguchi等[16]則觀察到HSOS患者血清VEGF水平顯著升高。我們的研究表明,MCT顯著增加了HHSEC細胞中MMP-9和VEGFA的mRNA表達水平,與文獻報道結果一致。另外我們還發現MCT可降低HHSEC中NOS3的mRNA表達水平。內皮一氧化氮合酶(eNOS)是一種心血管系統中的重要酶,表達于SEC,可催化一氧化氮(NO)的生成,從而保護內皮細胞[17];在HSOS模型動物中,可見NO含量顯著下降[18]。我們的研究表明,MCT降低了NOS3的mRNA表達水平,這可能會減少NO的產生,從而進一步損傷內皮細胞加劇毒性。
綜上所述,MCT 經HHSEC內代謝酶代謝活化,同時消耗細胞內GSH破壞肝臟抗氧化系統。MCT代謝活化一方面激活了HHSEC凋亡,另一方面影響內皮及血管系統等進一步加劇HHSEC損傷。本研究探討了MCT對HHSEC的體外細胞毒性及其毒性機制,為PAs所致臨床HSOS疾病的研究提供了理論依據。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
