生物炭與菌肥配施對元寶楓育苗基質性質及幼苗生長的影響
2020-06-21 15:35:06白雪李小英李俊龍
江蘇農業科學 2020年9期
白雪 李小英 李俊龍
關鍵詞:生物炭;菌肥;基質性質;元寶楓;生理生長特性
生物炭(biochar)通常指農林廢棄物等生物質在缺氧條件下不完全燃燒產生的富碳產物,炭化后具有多微孔結構和較大的比表面積,施入土壤后能夠降低土壤容重,改善土壤結構性,增加土壤中的有效養分,提升肥料利用率[1-2]。相關研究表明,單施生物炭能夠促進作物生長或增產,而生物質炭與有機肥料配施能夠更有效地提高土壤肥力,且不同配施方式生物質炭對土壤改良作用不盡相同,對作物的生長發育影響也不同[3-4]。菌肥是一類含有活性物質且可以獲得特定肥料效應的生物制品,菌肥中含有大量有效活菌,可以優化土壤微生物種群結構,提高土壤微生物量碳、氮含量,增強土壤酶活性,促進植物對養分的吸收[5-8]。Rong等研究認為,生物炭的多孔結構有益于土壤微生物存活,將生物炭與肥料配施,可以起到緩釋土壤養分的作用,而菌肥的作用機制是依靠大量的有益微生物來改善土壤的微生物群落結構,從而提高土壤養分利用效率[9]。表明生物炭和菌肥可以形成互補機制,作為菌肥的增效載體共同促進植株的生長發育。目前,大多數研究者利用生物炭的堿性來改良酸化土壤或中性土壤,將生物炭運用到育苗基質改良中的研究還鮮有報道。
元寶楓(Acer truncatum Bunge)是槭樹科(Aceraceae)槭樹屬植物,其根系較深,具有耐旱、抗寒、抗風害等特點,是兼有觀賞價值和經濟價值的優良樹種;其種仁含油率極高,且油中富含神經酸;其葉及種皮中也含有大量有益的生物活性物質,如黃酮、綠原酸、多糖、維生素、氨基酸、超氧化物岐化酶(SOD)等;其木材是制作家具的優質材料,種仁油渣可綜合利用,樹皮及果葉都可提取栲膠[10-11]。現階段,元寶楓籽油、楓葉生物活性物質作為保健食品和醫藥方面的新型優質原料,經常面臨原料緊缺的狀況,因此,須要加快培育優質元寶楓苗木,而元寶楓幼苗的培育須要重視苗木培育基質,普通的苗木基質已經不能滿足培育優質壯苗的需要。本研究利用生物炭的性能結合菌肥試驗,研究不同用量菌肥與生物炭結合對基質pH值、基質養分含量、基質酶活性及元寶楓幼苗生長的影響,以期獲取改良基質,為培育優質元寶楓苗木提供理論依據。
1 材料與方法
1.1 研究區概況
試驗地點為云南磷化集團有限公司,位于云南省昆明市晉寧區(地理位置為24°68′N,102°58′E),屬亞熱帶高原季風氣候,年均降水量為904.4 mm,年均氣溫為15 ℃。
1.2 試驗材料
供試土壤取自云南省昆明市晉寧區云南磷化集團有限公司大田,土壤類型為紅壤,土樣取回后自然晾干,粉碎,過2 mm篩后備用,并進行背景值分析,供試土壤基本性狀見表1;試驗選用長勢一致的一年生元寶楓幼苗;試驗用花盆直徑為25 cm,高 30 cm,每個盆中均裝入相同配比基質(珍珠巖、泥炭土、紅壤體積比為7.5 ∶ 0.5 ∶ 2.0)。基質材料為春城牌膨脹珍珠巖,丹麥品氏泥炭土;生物炭為永安優選炭業有限公司生產的椰殼炭(氮質量分數<0.5%,碳質量分數=63.37%,氫質量分數=35%),菌肥為山東綠隴生物科技有限公司生產的菌肥(有效活菌數含量≥200億CFU/g)。
1.3 試驗設計
每個花盆中均加入等量(1.3 kg)基質,按照不同處理,將生物炭、菌肥與基質混勻待用。其中生物炭(A)設置0、20 g/盆,菌肥(B)設置4個梯度,分別為0、10、20、30 g/盆,共3種處理(無添加CK、單施生物炭、生物炭與菌肥配施),即5種處理組合,分別為CK、AB0、AB1、AB2、AB3,生物炭與菌肥施用量見表2,每個處理組合重復10次。
1.4 測定內容與方法
1.4.1 基質性質測定 于苗木移栽90 d后對上述基質的性質進行測定[12]。有機質含量采用重鉻酸鉀-硫酸外加熱法測定;土壤全氮含量采用凱氏定氮法測定;土壤全磷含量、全鉀含量采用硫酸-高氯酸消煮法測定;土壤速效磷含量采用鹽酸-氟化銨浸提法測定;土壤速效鉀含量采用乙酸銨浸提-火焰光度計法測定;pH值用pH計測定;蔗糖酶活性采用3,5-二硝基水楊酸法測定;過氧化氫酶活性采用高錳酸鉀滴定法測定;脲酶活性采用鉬藍比色法測定。不同指標均重復測定3次。
1.4.2 生理指標測定 于2019年7月8日進行植物生理指標測定,每個處理抽取3株,選取植株自上而下的第3、第4張葉片進行凈光合速率(Pn)、胞間CO2濃度(Ci)、氣孔導度(Gs)、蒸騰速率(T)、葉綠素含量、葉片水分利用效率(WUE)的測定,其中凈光合速率、胞間CO2濃度、氣孔導度、蒸騰速率、葉片水分利用效率等用Li-6400XT便攜式光合作用測量系統進行測定,葉綠素含量用無水乙醇法測定[13]。
1.4.3 生長指標測定 元寶楓幼苗于2018年12月18日移栽,平均株高為30.46 cm,平均莖粗為 4.43 mm,從幼苗移栽日至2019年2月底期間幼苗均處于生長休眠期,2019年3月上旬才陸續發芽、發葉,2019年3月8日每個處理選取3株長勢均勻的幼苗測定株高、莖粗,試驗期間不進行任何施肥處理,僅定期澆水。
生長指標與生理指標同期測定,株高測從基質表面到頂芽底部的高度,用直尺(精度1 mm)測定;莖粗用游標卡尺(精度0.1 mm)測定;單株葉面積用Li-300A便攜式葉面積測量儀測定;根系體積用排液法測定;于2019年7月8日從每個處理中選取3株長勢一致的幼苗,對株高、莖粗進行復測,與2019年3月8日測定的幼苗株高、莖粗數值比較以獲得株高增長量和莖粗增長量等數據。
1.5 數據處理與分析
試驗數據用WPS 2010進行數據處理和圖表繪制,采用SPSS 22(IBM SPSS Statistic)軟件對數據進行統計分析。
2 結果與分析
2.1 不同處理育苗基質理化性質比較
從表3可以看出,物理性質方面,不同處理基質容重、總孔隙度與CK差異顯著。單施生物炭、生物炭與菌肥混施均可降低基質容重、提高基質總孔隙度,且AB3處理的效果最好,顯著降低了基質容重,提高了基質總孔隙度;與CK比較,單施生物炭容重降低29.6%,總孔隙度提高26.7%;生物炭與菌肥混施更好地降低了土壤容重,提高了土壤總孔隙度,與CK比較,其中AB3處理容重降低40.0%,總孔隙度提高34.5%。化學性質方面,不同處理與CK比較各項指標均差異明顯。有機碳含量、全氮含量、堿解氮含量、全磷含量、速效磷含量、全鉀含量、速效鉀含量均隨著菌肥施用量增加而呈增加的趨勢,其他處理各項化學性質指標均高于對照(除AB0處理基質速效磷含量外),其中菌肥高施用量處理(AB3)有機碳含量提高 38.5%,全氮含量提高89.1%,堿解氮含量提高 53.6%,全磷含量提高168%,速效磷含量提高 21.1%,AB0處理全鉀含量最高,比對照提高 18.9%,AB2處理速效鉀含量最高,相較于對照提高66.1%。
2.2 不同處理基質酶活性比較
從表4可以看出,不同處理基質的過氧化氫酶活性、脲酶活性、蔗糖酶活性變化情況。與CK比較,單施生物炭處理過氧化氫酶活性、脲酶活性分別降低15.79%、10.41%,而蔗糖酶活性提高1175%;生物炭與菌肥混施中高用量、中用量菌肥處理(AB2、AB3)的基質酶活性均顯著提高,過氧化氫酶活性分別提高42.11%、17.54%,脲酶活性分別提高136.20%、105.43%,蔗糖酶活性分別提高10470%、167.40%,低菌肥施用量與生物炭配施處理(AB1)僅僅提高了基質的脲酶活性;與單施生物炭相比較,生物炭與菌肥配施處理均提高了基質中酶活性。
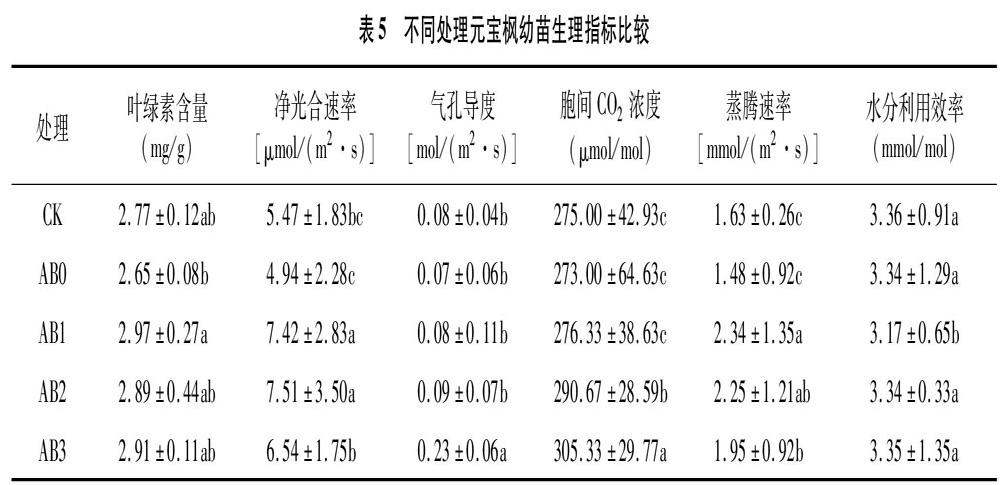
2.3 不同處理對元寶楓幼苗生理特性的影響
從表5可以看出,與CK比較,單施生物炭處理幼苗的葉綠素含量、Pn、Ci、Tr、WUE、Gs均降低;生物炭與菌肥配施處理幼苗的葉綠素含量、Pn、Gs、Ci、Tr均有所提高,但WUE降低,其中AB1處理葉綠素含量最高,提高 7.2%,AB2處理對Pn、Tr影響顯著,分別提高37.3%、38.0%,AB3處理對Gs、Ci影響顯著,分別提高187.5%、11.0%。與CK的各項生理指標比較,生物炭與菌肥配施處理對各項生理指標的影響明顯。
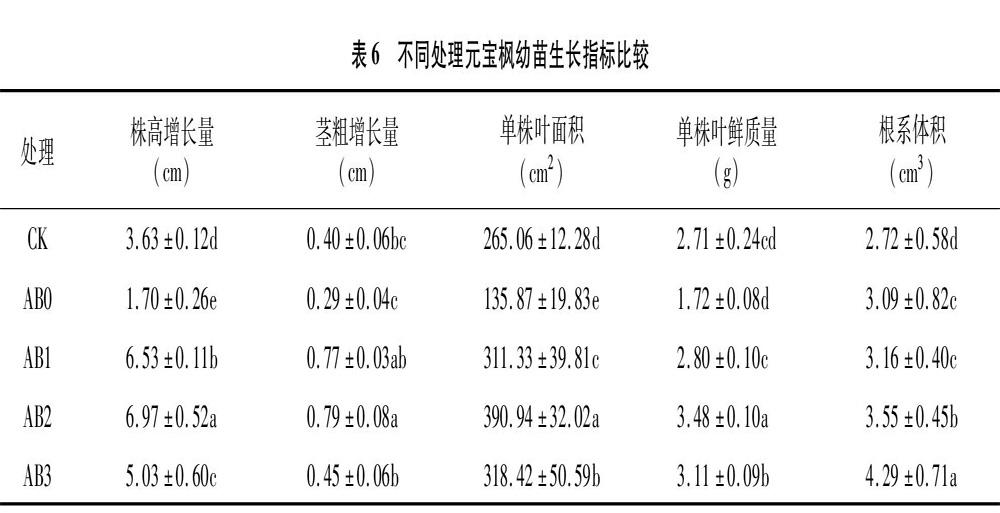
2.4 不同處理對元寶楓幼苗生長特性的影響
從表6可以看出,不同處理對元寶楓幼苗的株高增長量、莖粗增長量、單株葉鮮質量、單株葉面積、根系體積的影響。與CK相比,單施生物炭抑制了幼苗地上部分生長,株高增長量、莖粗增長量、單株葉鮮質量、單株葉面積均降低,分別降低5317%、27.50%、36.53%、48.74%,但根系體積提高;生物炭與菌肥混施處理幼苗株高增長量、莖粗增長量、單株葉鮮質量、單株葉面積、根系體積均提高,表明生物炭與菌肥混施能夠有效促進元寶楓幼苗的生長,其中AB2處理可以顯著提高幼苗株高增長量、莖粗增長量、單株葉鮮質量、單株葉面積,各項指標比CK分別提高92.0%、97.5%、28.4%、475%,AB3處理根系體積顯著增加,比CK增加57.57%。
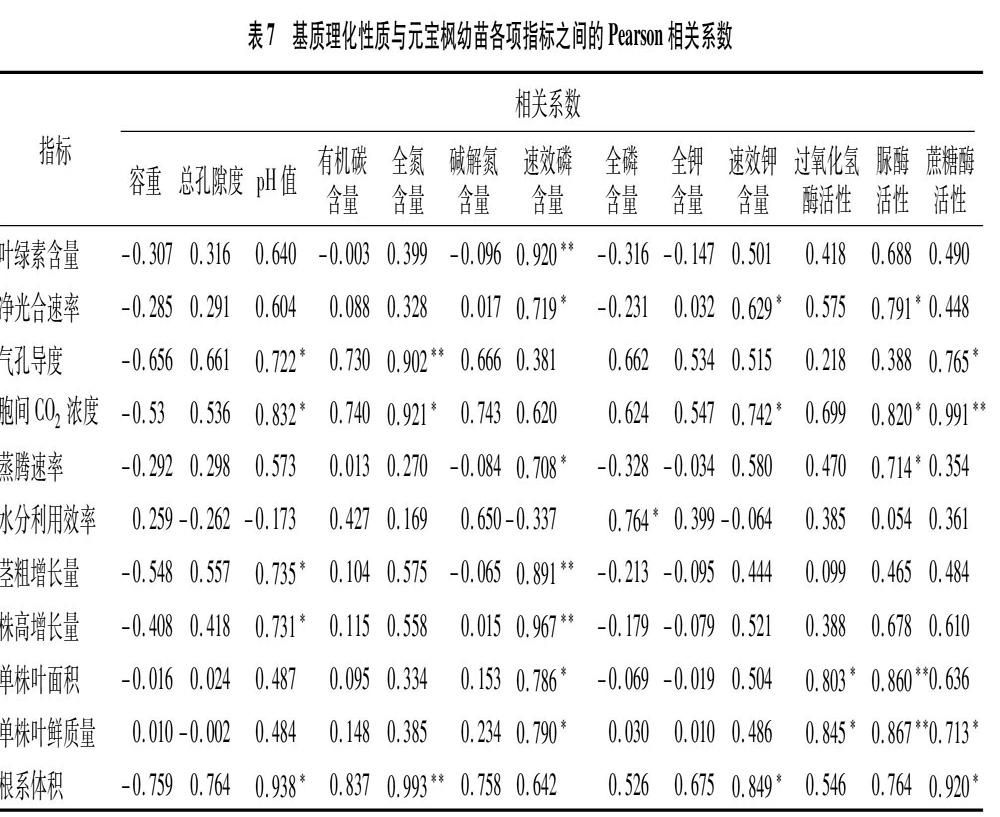
2.5 基質理化性質與幼苗生長指標相關性分析
基質理化性質與元寶楓幼苗各項生長指標之間的相關性分析結果見表7,總體上,基質容重與元寶楓幼苗各項生長指標(水分利用效率除外)呈負相關,基質總孔隙度與元寶楓幼苗生長各項指標(水分利用效率除外)呈正相關;pH值、有機碳含量、全氮含量、速效磷含量、速效鉀含量、酶活性與元寶楓幼苗生長各項指標(水分利用效率除外)呈正相關,其中基質pH值與幼苗Gs、Ci、莖粗增長量、株高增長量、根系體積呈顯著正相關(P<0.05);基質全氮含量與Ci呈顯著正相關(P<0.05),與Gs、根系體積呈極顯著正相關(P<0.01),相關系數分別為0.902、0.993;基質速效磷含量與幼苗Pn、Tr、單株葉面積、單株葉鮮質量呈顯著正相關(P<005),與葉綠素含量、莖粗增長量、株高增長量呈極顯著正相關(P<0.01),相關系數分別為0.920、0.891、0.967;基質全磷含量與WUE呈顯著正相關(P<0.05),與幼苗其他各項指標均無顯著相關性;基質速效鉀含量與Pn、Ci、根系體積呈顯著正相關(P<0.05);基質過氧化氫酶活性與單株葉面積、單株葉鮮質量呈顯著正相關(P<0.05);基質脲酶活性與Pn、Ci、Tr呈顯著正相關(P<0.05),與單株葉面積、單株葉鮮質量呈極顯著正相關(P<0.01),相關性系數分別為0.860、0.867;蔗糖酶活性與Gs、單株葉鮮質量、根系體積呈顯著正相關(P<0.05),與Ci呈極顯著正相關(P<0.01)。
3 討論與結論
3.1 添加生物炭與菌肥對基質理化性質的影響
生物炭、菌肥加入基質后各項性質會因土壤類型、水熱條件不同而產生差異,育苗基質的性質直接影響幼苗的栽培效果, 目前還未見作物栽培基質的標準化性狀參數[14]。相關研究表明,理想栽培基質容重為0.1~0.8 g/cm3,總孔隙度為54%~96%,pH值以弱酸性或中性為宜[15]。本試驗中,總體上基質的化學性質與幼苗的各項性質呈正相關,除CK外其他4種處理的容重及總孔隙度均在此范圍內,pH值均為弱酸性,單施生物炭或生物炭與菌肥配施處理均能顯著降低基質容重、增加基質孔隙度、降低基質酸性,對基質結構和物理性質起到了改良作用,與CK相比,AB0處理基質容重下降4522%,AB3處理基質總孔隙度提升34.48%;與CK比較,AB0處理基質的全氮含量、堿解氮含量、全鉀含量、速效鉀含量、全磷含量均提升,但速效磷含量降低30.60%,生物炭與菌肥配施處理基質的養分含量隨著菌肥施用量增加而增加,且AB3處理效果較好,說明生物炭、菌肥均能夠有效提升基質肥力,且生物炭與菌肥配施能夠減少基質中速效養分的流失。相關研究表明,植煙地施加生物炭與菌肥比單施菌肥能夠更好地降低土壤容重,增加土壤孔隙度,并能顯著提高土壤速效鉀及有機碳含量,在總體上提升土壤肥力[16-17]。本研究結果與之相似,說明基質的理化性質能夠直接影響幼苗的生長發育[18]。生物炭和菌肥配施使得生物炭的特性與菌肥形成了互補機制,更好地激發了二者之間對于基質養分的效用,起到了改良基質的作用。
3.2 添加生物炭與菌肥對基質酶活性的影響
土壤酶是土壤的組成部分,能夠反映土壤生物活性的高低,表征土壤養分轉化的快慢,可以作為土壤肥力的指標,脲酶活性可以表征基質中氮的轉化,蔗糖酶活性可表征基質中有機質含量、微生物數量及活動能力,過氧化氫酶活性可表征基質對土壤毒害作用的抵御能力[19-20]。相關研究中施用生物炭對土壤酶活性的影響結果不一,有提高[21],有降低[22],這可能與土壤、生物炭自身性質不同有關;本試驗中,各處理間的過氧化氫酶活性、脲酶活性大小關系分別表現為AB2>AB3>CK>AB1>AB0、AB2>AB3>AB1>CK>AB0,蔗糖酶活性大小關系表現為AB3>AB2>AB0>CK>AB1,生物炭與菌肥配施處理基質酶活性顯著提高,且酶活性與幼苗生理生長指標均呈正相關,說明基質中添加生物炭和菌肥可以有效改變基質微生物環境,酶活性增強使得基質中微生物活性增強,促進基質養分轉化,最終促進了幼苗的生長發育。相關研究表明,施加菌肥能有效提高土壤酶活性,能夠促進平邑甜茶幼苗的生長,且生物炭與菌肥配施較單施菌肥土壤酶活性增幅更大,能夠更好地提升平邑甜茶幼苗的生物量積累,促進幼苗根系發育[23-24]。本研究中生物炭添加菌肥處理可以促進元寶楓幼苗生長發育的研究結果與之相似。
3.3 添加生物炭與菌肥對幼苗生長、生理特性的影響
生物質炭化學性質穩定不易分解,但不能作為養分的直接來源,但生物質炭可以作為肥料的載體,吸附緩釋養分,提高肥料利用率,進而改善幼苗生長、生理特性[25]。光合作用是植物生長代謝最基本的物質和能量來源,凈光合速率的大小可以表征植株生長發育過程中生長代謝的水平,而葉綠素含量的高低很大程度上反映了植物的生長狀況和光合能力,氣孔導度、葉片水分利用效率是表征植物的耐受性[26],測定元寶楓幼苗的生長生理指標,可以綜合反映不同處理基質對幼苗生長的影響。本試驗中,單施生物炭處理在幼苗生長特性指標方面總體表現出抑制,黃曉麗等研究表明,生物炭能促進美國紅楓幼苗生長[27],這可能是由不同樹種自身生長發育特點、以及試驗區域的差異引起的。孫琪然等研究表明,生物炭與菌肥配施比單施生物炭、單施菌肥,能夠更好地提升平邑甜茶幼苗的生物量積累,促進幼苗根系發育[28]。與CK比較,單施生物炭處理幼苗的各項生長生理指標均降低,生物炭與菌肥配施處理(AB1、AB2、AB3)總體表現顯著促進幼苗生長發育,其中株高增長量、莖粗增長量大小關系表現為AB2>AB3>AB1,單株葉面積、單株葉鮮質量大小關系表現為處理AB2>AB1>AB3,根系體積大小關系為處理AB3>AB2>AB1,總體上高菌肥施用量對幼苗地上部分表現出生長抑制,對地下部分表現出生長促進。生物炭與菌肥配施的葉綠素含量、Pn、Gs、Ci、Tr均有所提高,其中AB1處理葉綠素含量最高,比CK提高7.2%,AB2的Pn、Tr較高,分別比CK提高37.3%、38.0%,AB3的Gs、Ci最高,分別比CK提高187.5%、11.0%。表明基質的養分轉化吸收過程復雜,不僅與基質養分含量和不同養分之間的相互作用密切相關,還與幼苗的生理生長特性有關。
單施生物炭、生物炭和菌肥配施均可降低基質容重,增加基質總孔隙度,改善基質酸堿性,提高基質養分含量,總體上基質養分含量隨菌肥施用量增加而增加,其中AB3處理對基質有效養分提升效果最優,堿解氮含量、速效磷含量、速效鉀含量,與CK比較分別提高53.57%、21.07%、64.59%。總體上,基質容重與元寶楓幼苗各項指標呈負相關,基質總孔隙度與元寶楓幼苗各項指標呈正相關;pH值、有機碳含量、全氮含量、堿解氮含量、速效磷含量、速效鉀含量、酶活性與元寶楓幼苗大多數指標呈正相關,表明基質理化性質直接影響了幼苗的生長發育,適宜的基質能夠促進幼苗的生長。綜合考慮經濟效益,AB2處理基質可用于改良基質及培育元寶楓優質壯苗,并可作為優良基質在生產上推廣應用。
從本試驗結果可知,生物炭與菌肥合理配施是提升基質肥力、培育壯苗的關鍵。但本研究只是針對現有基質結合生物炭及菌肥的性能對元寶楓幼苗基質的改良,試驗設置還比較單一,如要獲得其他苗木的改良培育基質可在本研究的基礎上改變生物炭及菌肥施用梯度設置,將可獲得更多可用的苗木基質改良配方。
參考文獻:
[1]孟繁昊,高聚林,于曉芳,等. 生物炭配施氮肥改善表層土壤生物化學性狀研究[J]. 植物營養與肥料學報,2018,24(5):1214-1226.
[2]Sohi S P,Krull E,Lopez-Capel E,et al.A review of biochar andits use and function in soil[J]. Advances in Agronomy,2010,105(1):47-82.
[3]文 靜,楊丹丹,林啟美,等. 生物質炭復混肥對土壤肥力與玉米和大豆生物量的影響[J]. 中國土壤與肥料,2015(3):74-78.
[4]劉祥義,付 惠,邱宗海. 云南元寶楓葉營養評價[J]. 天然產物研究與開發,2003,15(3):222-223,226.
[5]李 樂,孫 海,劉政波,等. 微生物肥料的作用、機理及發展方向[J]. 東北農業科學,2016,41(4):63-69.
[6]盧培娜,劉景輝,趙寶平,等. 菌肥對鹽堿地土壤特性及燕麥根系分泌物的影響[J]. 作物雜志,2017(5):85-92.
[7]張曼玉,高 婷,吳永波,等. 生物炭對貴州喀斯特山地石漠化土壤理化性質和構樹幼苗生長特性的影響[J]. 江蘇農業科學,2019,47(12):177-181.
[8]Lu Y,Yu M J,Xing L,et al. Combined application of biochar and nitrogen fertilizer benefits nitrogen retention in the rhizosphere of soybean by increasing microbial biomass but not altering microbial community structure[J]. Science of the Total Environment,2018,640-641:1221-1230.
[9]Rong Q L,Li R N,Huang S W,et al. Soil microbial characteristics and yield response to partial substitution of chemical fertilizer with organic amendments in greenhouse vegetable production[J]. Journal of Integrative Agriculture,2018,17(6):1432-1444.
[10]吳衛中,李 珺,許 疌. 元寶楓化學成分的研究概況[J]. 中國藥事,2008,22(7):603-609.
[11]王 典,張 祥,姜存倉,等. 生物質炭改良土壤及對作物效應的研究進展[J]. 中國生態農業學報,2012,20(8):963-967.
[12]鮑士旦. 土壤農化分析[M]. 北京:中國農業出版社,2000.
[13]高俊鳳. 植物生理學實驗指導[M]. 北京:高等教育出版社,2006.
[14]李 婭,李恩良,毛云玲,等. 不同育苗基質對長蕊甜菜樹苗木生長的影響[J]. 東北林業大學學報,2019,47(3):8-11,21.
[15]呂英忠,習玉森,王新平,等. 不同基質對蘋果幼樹生長發育及葉片生理特性的影響[J]. 中國農學通報,2018,34(34):47-54.
[16]李 影,李 斌,柳東陽,等. 生物炭配施菌肥對植煙土壤養分和可溶性有機碳氮光譜特征的影響[J]. 華北農學報,2018,33(6):227-234.
[17]李 影. 有機菌肥和生物炭配施對豫中烤煙生長、品質及植煙土壤性質的影響[D]. 鄭州:河南農業大學,2018:9-12.
[18]He L L,Zhong Z K,Yang H M. Effects on soil quality of biochar and straw amendment in conjunction with chemical fertilizers[J]. Journal of Integrative Agriculture,2017,16(3):704-712.
[19]胡慧蓉,田 昆. 土壤學實驗指導教程[M]. 北京:中國林業出版社,2012.
[20]李新宇,孟 康,李小英,等. 生物炭對元謀燥紅壤土壤肥力與番茄生長的影響[J]. 西部林業科學,2019,48(2):114-120.
[21]鄭慧芬,吳紅慧,翁伯琦,等. 施用生物炭提高酸性紅壤茶園土壤的微生物特征及酶活性[J]. 中國土壤與肥料,2019(2):68-74.
[22]許云翔,何莉莉,劉玉學,等. 施用生物炭6年后對稻田土壤酶活性及肥力的影響[J]. 應用生態學報,2019,30(4):1110-1118.
[23]王義坤,孫琪然,段亞楠,等. 三種菌肥對蘋果連作土壤環境及平邑甜茶幼苗生長的影響[J]. 植物營養與肥料學報,2019,25(4):630-638.
[24]趙曉軍,李 麗,張 璇,等. 生物炭與微生物菌劑配施對土壤生物和化學特性的影響[J]. 安徽農業科學,2018,46(25):109-112.
[25]嚴陶韜,丁子菊,朱 倩,等. 生物質炭對黃棕壤理化性質及龍腦樟幼苗生長的影響[J]. 土壤,2018,50(4):681-686.
[26]陳媛媛,江 波,王效科,等. 元寶楓幼苗生長和光合特性對硬化地表的響應[J]. 生態學雜志,2016,35(12):3258-3265.
[27]黃曉麗,葛根塔娜,梅 梅,等. 生物炭對美國紅楓幼苗生長及葉色變化的影響[J]. 沈陽農業大學學報,2017,48(5):530-536.
[28]孫琪然,徐 燕,相 立,等. 生物炭和菌肥的混合使用對蘋果園連作土壤環境及平邑甜茶幼苗生理指標的影響[J]. 中國農學通報,2017,33(8):52-59.李新蕾,李葉芳,關文靈. PEG模擬干旱脅迫對大葉醉魚草種子萌發及幼苗生理特性的影響[J].
