溫度調(diào)控下枯草芽孢桿菌配合酵母對褶皺臂尾輪蟲種群增長的影響
2020-06-21 15:35:06肖佳華李茹夢邵力
江蘇農(nóng)業(yè)科學(xué) 2020年9期
肖佳華 李茹夢 邵力
關(guān)鍵詞:枯草芽孢桿菌;褶皺臂尾輪蟲;酵母;溫度調(diào)控;種群增長
褶皺臂尾輪蟲(Brachionus plicatilis)是海水魚、蝦、蟹等水產(chǎn)幼苗生長發(fā)育期間不可或缺的餌料生物,“育好苗先培好蟲”這句行話說明輪蟲培育是育苗產(chǎn)業(yè)上不可缺少的關(guān)鍵一環(huán)。褶皺臂尾輪蟲食性廣泛,生產(chǎn)上主要以投喂單胞藻為主,但土池生產(chǎn)中單胞藻的接種和培育受天氣環(huán)境等不可控因素的影響較大,工廠化生產(chǎn)中單胞藻的接種和培育則受場地限制等因素影響較大,因此單胞藻產(chǎn)量極其不穩(wěn)定,池塘內(nèi)常常因?yàn)槌霈F(xiàn)藻類的快速演替,從而導(dǎo)致輪蟲餌料短缺的問題,嚴(yán)重影響育苗產(chǎn)業(yè)的經(jīng)濟(jì)效益。在單胞藻缺乏時,酵母是被廣泛運(yùn)用的輪蟲基礎(chǔ)餌料,20世紀(jì)60年代Hirata發(fā)現(xiàn)可以用面包酵母培養(yǎng)海水輪蟲[1],20世紀(jì)70年代海水小球藻與面包酵母培養(yǎng)輪蟲的模式被廣泛推廣[2],相比小球藻,酵母生產(chǎn)廠家眾多,可獲得性好,發(fā)酵條件簡單,成本低廉,可在短時間內(nèi)替代單胞藻作為輪蟲的主要餌料,酵母已經(jīng)被證實(shí)可用于輪蟲高密度生產(chǎn)養(yǎng)殖,可保證在單胞藻供應(yīng)不足情況下維持輪蟲的存活和生長。但相關(guān)研究已經(jīng)證實(shí)單獨(dú)用酵母培養(yǎng)的輪蟲,易產(chǎn)生營養(yǎng)缺乏癥狀,如今輪蟲營養(yǎng)強(qiáng)化的研究涉及到藻類和酵母搭配光合細(xì)菌、魚油、維生素B12和維生素C等[3-8],甘松永等用酵母與枯草芽孢桿菌作為輪蟲餌料進(jìn)行高密度培養(yǎng),結(jié)果證實(shí)酵母和芽孢桿菌可用作餌料培養(yǎng)[9]。
本試驗(yàn)針對枯草芽孢桿菌生長特性及配合酵母投喂后對輪蟲種群生殖的影響作進(jìn)一步研究,以期獲得枯草芽孢桿菌與酵母搭配的最佳添加濃度和控制溫度,盡可能為實(shí)際生產(chǎn)當(dāng)中所遇到的問題提供數(shù)據(jù)支持,也為輪蟲營養(yǎng)強(qiáng)化提供新的思路。
1 材料與方法
1.1 試驗(yàn)材料的準(zhǔn)備
本試驗(yàn)中使用的試驗(yàn)菌株為枯草芽孢桿菌粉活化后分離篩選得到,經(jīng)生物性狀和培養(yǎng)試驗(yàn)鑒定為合格的水產(chǎn)養(yǎng)殖專用枯草芽孢桿菌菌株,枯草芽孢桿菌菌粉購自濟(jì)寧金山生物工程有限公司。
本試驗(yàn)使用的褶皺臂尾輪蟲為鹽城射陽海水育苗基地輪蟲養(yǎng)殖池塘內(nèi)取塘口底泥休眠卵孵化而來,試驗(yàn)室孵化后進(jìn)行單個體純化培養(yǎng),純化培養(yǎng)后收集休眠卵以保種,輪蟲培養(yǎng)液采用人工海水配方,并每天更換培養(yǎng)液,培養(yǎng)條件:光照度約為 4 000 lx,晝夜比為 16 h ∶ 8 h。
種子培養(yǎng)基:1.50%葡萄糖、1.50%蛋白胨、015%牛肉膏,pH值7.0~7.2,121 ℃滅菌30 min。
液體培養(yǎng)基:1.50%葡萄糖、1.50%蛋白胨、030% K2HPO3、0.10% 30.8 mg/L MnSO4、0.70% CaCO3、0.05% MgSO4·7H2O、0.01% FeCl3,pH值為7.0~7.2,121 ℃滅菌30 min。
斜面培養(yǎng)基:將15 g營養(yǎng)瓊脂溶入1 000 mL液體培養(yǎng)基中,121 ℃滅菌30 min,趁熱倒入已滅菌的培養(yǎng)皿,4 ℃冷藏。
酵母輪蟲基礎(chǔ)食物懸液:取干酵母加入濃度為1.5%葡萄糖培養(yǎng)基,溫度為25 ℃,活化30 min后,以22 ℃、5 500 r/min離心去除葡萄糖培養(yǎng)基,加入人工海水配成濃度為6×106個/mL酵母液備用。高活性干酵母購自安琪酵母股份有限公司。
1.2 試驗(yàn)方法
1.2.1 枯草芽孢桿菌活菌生長曲線的測定方法 在無菌操作臺下,將4 ℃保存的斜面培養(yǎng)基菌株接入已滅菌的新斜面培養(yǎng)基中,置于菌種培養(yǎng)箱 37 ℃ 活化24 h后接種到100 mL種子培養(yǎng)基中,37 ℃,200 r/min,搖床振蕩培養(yǎng)24 h。將培養(yǎng)后的種子液按7%的接種量接入已滅菌的100 mL液體培養(yǎng)基中,37 ℃、200 r/min搖床振蕩培養(yǎng)枯草芽孢桿菌母液,培養(yǎng)0、1、3、5、7、10、12、15、20、24、28 h時對枯草芽孢桿菌進(jìn)行計數(shù)。測定各時間點(diǎn)活菌數(shù)量時用1 mL無菌吸管吸取1 mL母液的菌懸液,加入9 mL無菌人工海水混勻成1 ∶ 10稀釋比例的菌懸液,這樣依次稀釋,直至得到1 ∶ 105、1 ∶ 106、1 ∶ 107、1 ∶ 108、1 ∶ 109、1 ∶ 1010等濃度,每次稀釋更換無菌吸管,以上每個濃度設(shè)置3次重復(fù)。向每個滅菌培養(yǎng)皿中倒入約10 mL已滅菌的液體培養(yǎng)基,混合均勻,待凝固后備用,用 1 mL 無菌吸管分別吸取不同稀釋濃度活菌懸液 0.1 mL,加至已凝固好的培養(yǎng)基中,用涂布棒涂勻,在無菌操作臺中靜置 30 min 后,倒置于37 ℃的恒溫培養(yǎng)箱內(nèi)培養(yǎng)18~24 h,對同一梯度平行平板菌落數(shù)進(jìn)行統(tǒng)計,計算平均值,根據(jù)稀釋倍數(shù)和取樣接種量換算樣品中活菌含量。
1.2.2 輪蟲種群動態(tài)研究方法 取冷藏保存的輪蟲休眠卵,放入海水培養(yǎng)液中,在自然光照、25 ℃條件下進(jìn)行孵化,并于16~24 h內(nèi)觀察發(fā)育狀況,發(fā)育形成穩(wěn)定的群體后,挑取出日齡4 h以內(nèi)的輪蟲幼體置于一次性細(xì)胞培養(yǎng)板(6孔)中,每孔加入 5 mL 試驗(yàn)溶液,每孔5只輪蟲幼體,每組設(shè)置6個平行;在枯草芽孢桿菌剛進(jìn)入對數(shù)生長期的時間點(diǎn)聯(lián)合酵母配成試驗(yàn)溶液,根據(jù)預(yù)試驗(yàn)結(jié)果,當(dāng)枯草芽孢桿菌的濃度超過1 000 mg/L時對輪蟲生長開始有抑制作用,設(shè)置枯草芽孢桿菌的濃度梯度為0(對照組)、100、200、300、500、800 mg/L,設(shè)置培養(yǎng)箱光照度為 4 000 lx,晝夜比為16 h ∶ 8 h,溫度分別設(shè)置為 20、25、30 ℃。每次間隔24 h鏡檢并更換培養(yǎng)液,連續(xù)觀察10 d,對輪蟲個體數(shù)進(jìn)行計數(shù),統(tǒng)計輪蟲種群數(shù)量、日種群增長率以及種群密度等。
1.3 數(shù)據(jù)處理
種群密度=種群總數(shù)/培養(yǎng)體積。種群密度單位為ind./mL。
根據(jù)收集的試驗(yàn)數(shù)據(jù),采用Excel作初步處理,再用Sigma Plot 11.0軟件進(jìn)行處理。根據(jù)試驗(yàn)設(shè)計,對數(shù)據(jù)進(jìn)行單因素方差分析(one-way ANOVA),并進(jìn)行多重比較,結(jié)果用平均值±標(biāo)準(zhǔn)差的形式表示。
2 結(jié)果與分析
2.1 枯草芽孢桿菌生長曲線測定
在培養(yǎng)0、1、3、5、7、10、12、15、20、24、28 h時對枯草芽孢桿菌活菌數(shù)進(jìn)行記錄,以活菌數(shù)量的對數(shù)作縱坐標(biāo),以培養(yǎng)時間作橫坐標(biāo),繪制活性菌株生長曲線,由圖1可知,枯草芽孢桿菌菌株從種子培養(yǎng)基接入液體培養(yǎng)基后經(jīng)歷了4個生長時期,分別為延遲期、對數(shù)生長期、穩(wěn)定期和衰退期,0~4 h為延遲期,4~10 h進(jìn)入對數(shù)增長期,活菌的數(shù)量呈指數(shù)型增長,峰值約達(dá)到1.1×1010CFU/mL后進(jìn)入穩(wěn)定期,穩(wěn)定期為10~24 h,活菌總體數(shù)量基本保持不變,24 h后進(jìn)入衰退期,活菌數(shù)量下降。因此,活化菌的時間應(yīng)控制在4 h左右,此時菌株剛進(jìn)入對數(shù)生長期,細(xì)菌形態(tài)典型、生物活性強(qiáng)。
2.2 20 ℃下不同濃度枯草芽孢桿菌聯(lián)合酵母對輪蟲種群的影響
2.2.1 20 ℃條件下輪蟲種群動態(tài) 20 ℃時,褶皺臂尾輪蟲在不同濃度枯草芽孢桿菌中種群動態(tài)變化
見圖2,輪蟲種群數(shù)量隨枯草芽孢桿菌的濃度升高先上升再降低。對照組最大種群容納量為 324 ind.,100、200、300、500、800 mg/L枯草芽孢桿菌處理組最大種群容納量分別為343、369、375、402、356 ind.。在種群起始密度均為1.0 ind./mL的情況下,除 300 mg/L 枯草芽孢桿菌處理組在第10天才到達(dá)種群數(shù)量高峰外,其他各處理組和對照組均在第9天達(dá)到種群數(shù)量高峰,之后種群數(shù)量呈下降趨勢。
2.2.2 20 ℃條件下輪蟲種群增長率 褶皺臂尾輪蟲在20 ℃條件下不同濃度枯草芽孢桿菌日種群增長率見圖3,試驗(yàn)組種群增長率始終高于對照組,500 mg/L枯草芽孢桿菌試驗(yàn)處理組在第5天達(dá)到各試驗(yàn)組中最大種群增長率,為16.3%,明顯高于對照組,各組種群增長率均隨時間先上升后下降。對照組在第5天達(dá)到峰值,處理組種群增長率峰值出現(xiàn)時間均與對照組一致。
2.2.3 20 ℃條件下輪蟲種群密度 20 ℃時,輪蟲在不同濃度枯草芽孢桿菌中種群密度變化見圖4。第2天,對照組與各試驗(yàn)處理組種群密度差異較小。第3天,各處理組與對照組相比種群密度顯著增大(P<0.05)。第9天,100、200、800 mg/L處理組與對照組相比,雖然種群密度增大,但差異不顯著(P>0.05)。第10天,種群密度普遍下降,500 mg/L 組種群密度較第9天減小,但與對照組差異保持顯著(P<0.05)。總體來看,在20 ℃條件下,除 300 mg/L 濃度之外,其他濃度組種群密度隨時間先增大后減小。
2.3 25 ℃條件下不同濃度枯草芽孢桿菌聯(lián)合酵母對輪蟲種群的影響
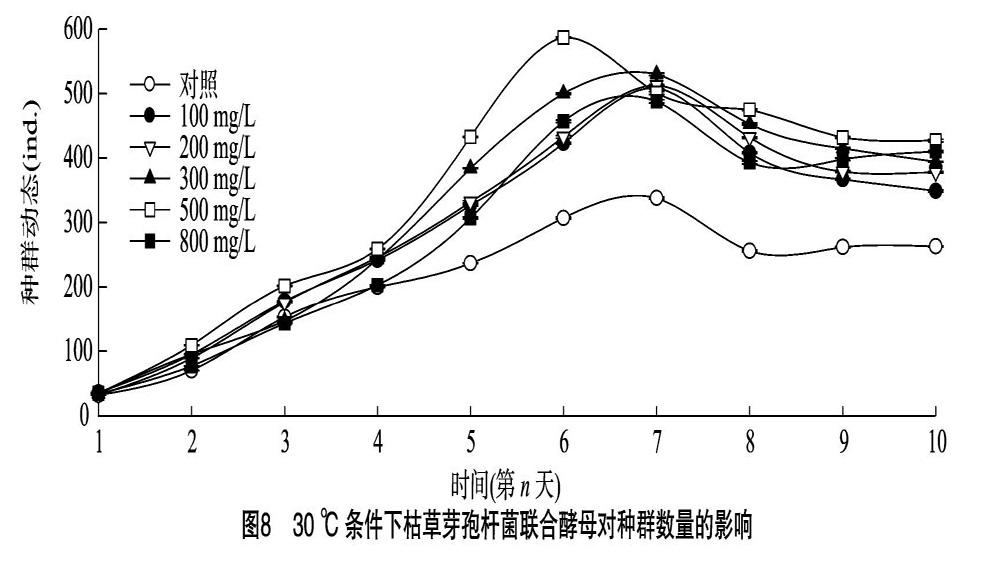
2.3.1 25 ℃條件下輪蟲種群動態(tài) 25 ℃條件下對照組和試驗(yàn)組最大種群容納量見圖5,對照組最大種群容納量達(dá)到333 ind.,100、200、300、500、800 mg/L枯草芽孢桿菌處理組的最大種群容納量分別達(dá)到603、647、723、842、639 ind.。試驗(yàn)周期內(nèi)各濃度處理組種群最大容量與對照空白組差異顯著(P<0.05),500 mg/L組最大容納量是空白對照組的2.53倍。除200 mg/L組之外,對照組及其他試驗(yàn)處理組到達(dá)峰值的時間相同,均在試驗(yàn)第9天。
2.3.2 25 ℃條件下輪蟲種群增長率 25 ℃條件下輪蟲種群增長率見圖6,對照組種群增長率隨時間延長呈先快速上升后緩慢下降的趨勢。100、200、300、800 mg/L處理組種群增長趨勢同對照組,呈先快速增長再緩慢下降的趨勢,4個濃度處理組最大種群增長率均在第3天出現(xiàn),較對照組提前1 d。第1天至第3天,800 mg/L組種群增長率最高,從第4天開始,500 mg/L種群增長率超越800 mg/L處理組,并保持緩慢增長趨勢,在第7天到達(dá)最大種群增長率后開始下降,在第10天,試驗(yàn)處理組與對照組之間差異顯著(P<0.05),但各試驗(yàn)組之間種群增長率差異不顯著(P>0.05)。總體而言,25 ℃時添加枯草芽孢桿菌后,種群增長率在試驗(yàn)周期內(nèi)始終高于空白對照組,在所有試驗(yàn)處理組中綜合種群增長率最大的是 500 mg/L 處理組,增速最快的是 800 mg/L 處理組。
2.3.3 25 ℃條件下輪蟲種群密度 由圖7可知,25 ℃時試驗(yàn)第2天,對照組輪蟲種群密度為(1.2±0.3) ind./mL,與100、200、300、500 mg/L處理組之間差異并不顯著(P>0.05),但此時對照組與最高濃度組之間種群密度差異顯著(P<0.05),最高濃度(800 mg/L)組種群密度達(dá)到(1.9±0.4) ind./mL,均大于對照組及其他處理組。試驗(yàn)第3天,對照組種群密度為(2.1±0.7) ind./mL,100、200、300、500、800 mg/L處理組種群密度分別為(3.1±0.7)、(2.8±0.5)、(3.3±0.6)、(3.0±0.6)、(3.8±0.7) ind./mL,各試驗(yàn)組均顯著高于對照組(P<0.05)。試驗(yàn)第9天,500 mg/L處理組種群密度達(dá)到峰值,為(28.1±0.4) ind./mL,為對照組濃度的2.55倍,300 mg/L處理組種群密度達(dá)到(24.1±2.3) ind./mL,是對照組的2.19倍,此時最高濃度組的種群密度與100、200 mg/L處理組差異不顯著(P>0.05),分別為(21.0±2.0)、(19.9±2.6)、(21.5±2.3) ind./mL,分別較對照組提高93.6%、82.7%、94.5%。試驗(yàn)第10天,各試驗(yàn)組種群密度保持穩(wěn)定, 增速降低。總體而言,在25 ℃溫度條件下,最適添加濃度控制在300~500 mg/L區(qū)間內(nèi)最佳。
2.4 30 ℃時不同濃度枯草芽孢桿菌聯(lián)合酵母對輪蟲密度的影響
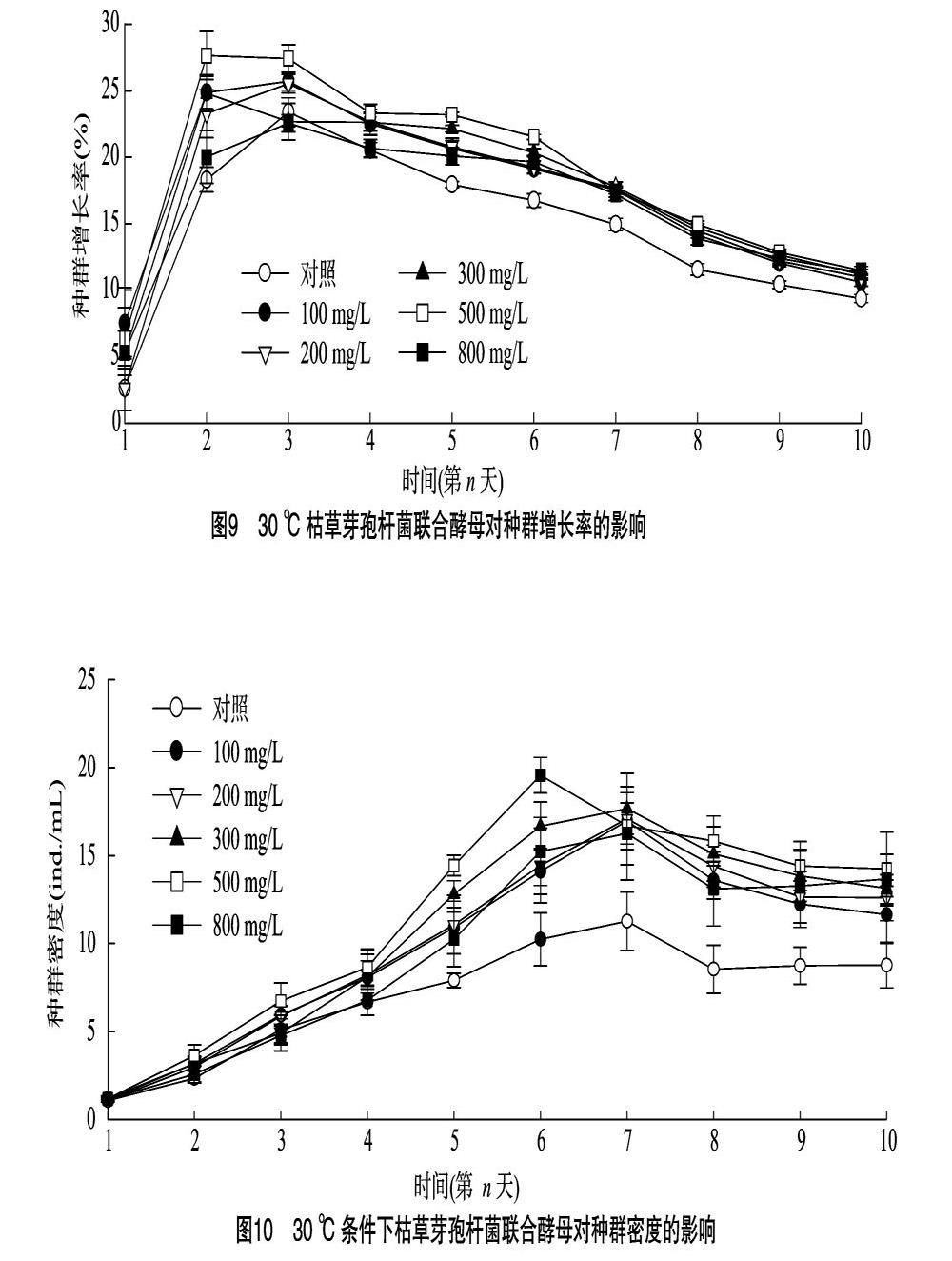
2.4.1 30 ℃條件下輪蟲種群動態(tài) 由圖8可知,30 ℃時, 對照組與100、200、300、500、 800 mg/L處理組的最大種群容納量分別為263、509、513、530、587、488 ind.,各處理組與對照組差異顯著(P<005)。最大種群容納量出現(xiàn)在500 mg/L處理組,其容納量是對照組的2.23倍。試驗(yàn)組容納量隨枯草芽孢桿菌濃度升高呈先升高后下降趨勢,第10天,800 mg/L試驗(yàn)組與100 mg/L試驗(yàn)組差異顯著(P<0.05)。100、200、300、800 mg/L處理組及對照組到達(dá)峰值的時間相同,均在試驗(yàn)第7天,500 mg/L處理組到達(dá)峰值的時間為第6天,較其他處理組提前1 d達(dá)到容納量峰值。
2.4.2 30 ℃條件下輪蟲種群增長率 由圖9可知,30 ℃條件下,所有組別種群增長率均隨時間延長呈先快速上升后下降。其中300、500 mg/L處理組最大種群增長率出現(xiàn)在第2天,較對照組提前 1 d,其中500 mg/L處理組種群增長率高于 300 mg/L 處理組。對照組與100、200、500 mg/L 處理組的最大種群增長率出現(xiàn)在第3天。在第3天時,800、300 mg/L處理組種群增長率低于空白對照組,其他試驗(yàn)組均高于空白對照組。種群增長率最大的是500 mg/L處理組。
2.4.3 30 ℃條件下輪蟲種群密度 由圖10可知,30 ℃時,單因素方差分析結(jié)果表明,在種群起始密度均等的情況下,第2天時對照組與100、200、300、500 mg/L處理組差異顯著(P<0.05),與800 mg/L處理組差異不顯著,各處理組間種群密度無顯著差異(P>0.05),此時500 mg/L處理組種群密度為(3.6±0.6) ind./mL,較對照組種群密度增加565%。第3天時,500 mg/L處理組與100、200 mg/L 處理組之間差異不顯著(P>0.05),與空白對照組差異顯著(P<0.05), 300、800 mg/L試驗(yàn)組保持緩慢增長,與空白對照組差異不顯著,不具有統(tǒng)計學(xué)意義(P>0.05)。第4天時,對照組除與 800 mg/L 對照組差異不顯著(P>0.05)外,與100、200、300、500 mg/L處理組均差異顯著。在試驗(yàn)第6天時,500 mg/L 試驗(yàn)組種群密度達(dá)到所有處理組中最大值,為(19.6±1.0) ind./mL,是此時空白對照組的1.96倍,在第6天后呈下降態(tài)勢,其他濃度(100、200、300、800 mg/L)組在第7天達(dá)到各自的最大種群密度,分別為(16.9±1.2)、(17.1±0.9)、(17.6±2.0)、(16.2±2.7) ind./mL,較此時對照組種群密度(11.2±1.7) ind./mL 分別上升509%、52.7%、57.1%、44.6%。總體而言,添加枯草芽孢桿菌的處理組最大種群密度均高于空白對照組,在受試濃度范圍內(nèi),30 ℃條件下最適添加濃度范圍為300~500 mg/L。
3 討論
針對單胞藻生產(chǎn)不穩(wěn)定以及藻相突變引起的輪蟲餌料不足等問題,為短時間內(nèi)穩(wěn)定輪蟲產(chǎn)量,本試驗(yàn)對枯草芽孢桿菌配合酵母進(jìn)行研究。試驗(yàn)結(jié)果表明,在酵母作為主體餌料濃度一定的情況下,溫度和枯草芽孢桿菌濃度是影響褶皺臂尾輪蟲種群數(shù)量增長的主要因素。
3.1 枯草芽孢桿菌濃度對褶皺臂尾輪蟲的影響
枯草芽孢桿菌是水體中重要的微生態(tài)環(huán)境因子之一,在水產(chǎn)養(yǎng)殖生產(chǎn)活動中起著對水體生物殘留物及殘餌分解轉(zhuǎn)化的調(diào)節(jié)作用。用酵母培養(yǎng)輪蟲是向水體內(nèi)添加活酵母來保證輪蟲攝食,在生產(chǎn)中隨著酵母的投喂量增加,單位水體中的有機(jī)含量也逐漸增高,所以在輪蟲生長過程中用酵母液培養(yǎng)輪蟲,常常出現(xiàn)殘?jiān)^多水質(zhì)惡化的問題。酵母培養(yǎng)液中添加一定量枯草芽孢桿菌后,枯草芽孢桿菌可在水體中快速繁殖,同時產(chǎn)生大量的胞外酶類(如蛋白酶、脂肪酶、淀粉酶等),消耗水體中的生物殘留物,改善水質(zhì)降低化學(xué)需氧量和氨氮水平,同時分解出多種氨基酸和維生素類物質(zhì)等代謝產(chǎn)物,為輪蟲生長提供可利用的營養(yǎng)物質(zhì),對輪蟲種群增長起著促進(jìn)功能。本試驗(yàn)的結(jié)果顯示,輪蟲酵母培養(yǎng)液中添加枯草芽孢桿菌的效果表現(xiàn)明顯,各溫度條件下,處理組中輪蟲種群最大容納量和密度顯著高于空白對照組。溫度設(shè)置在20、25、30 ℃,在枯草芽孢桿菌設(shè)置的0~500 mg/L濃度區(qū)間內(nèi),褶皺臂尾輪蟲的種群數(shù)量增長效果明顯。在各個溫度節(jié)點(diǎn)之間,當(dāng)超過一定濃度的時候,設(shè)置過高的枯草芽孢桿菌濃度并不利于繼續(xù)擴(kuò)大種群容納量,尤其最高濃度 (800 mg/L) 的生長效果均明顯低于 500 mg/L 試驗(yàn)組,這與預(yù)試驗(yàn)得出當(dāng)枯草芽孢桿菌濃度超過 1 000 mg/L 時對褶皺臂尾輪蟲的生長是有明顯抑制作用的結(jié)論相符合。根據(jù)試驗(yàn)結(jié)果分析,在以酵母液為基礎(chǔ)培養(yǎng)液的情況下,活酵母菌進(jìn)入水體之后,在水體中的繼續(xù)擴(kuò)大繁殖消耗大量氧氣,當(dāng)過量的枯草芽孢桿菌加入酵母基礎(chǔ)培養(yǎng)液后則進(jìn)一步導(dǎo)致水體中氧氣不足,同時枯草芽孢桿菌因?yàn)榉N群繁殖優(yōu)勢取代酵母菌成為水體優(yōu)勢種,抑制酵母菌的生長,而枯草芽孢桿菌本身無法作為主體餌料滿足輪蟲生長營養(yǎng)需要,從而直接影響輪蟲攝食生長,嚴(yán)重時甚至導(dǎo)致輪蟲在短時間內(nèi)因濃度和饑餓雙重脅迫而快速死亡。在各溫度條件下,在以活酵母液為主體餌料的情況下,枯草芽孢桿菌最適添加濃度為500 mg/L。
3.2 溫度調(diào)控對褶皺臂尾輪蟲的影響
溫度是影響輪蟲種群動態(tài)的一個重要影響因子,不同溫度對影響輪蟲的種群動態(tài)產(chǎn)生顯著影響[10-12]。本試驗(yàn)結(jié)果顯示,隨著培養(yǎng)溫度的逐漸升高,輪蟲的最大種群增長率出現(xiàn)的時間縮短。說明,提高溫度可以有效地加快輪蟲的繁殖,在一定范圍內(nèi)繁殖速度與溫度呈正比,這與鄭樂云的試驗(yàn)結(jié)果[13]相符。鄭樂云對超小型輪蟲生長繁殖情況進(jìn)行研究,結(jié)果表明,在25~38 ℃溫度內(nèi)超小型輪蟲繁殖速度與溫度呈正相關(guān)關(guān)系,溫度越高,繁殖速度越快[13]。本試驗(yàn)結(jié)果表明,最大種群密度和最大容納量總體表現(xiàn)出隨著溫度的升高呈先增大后減小的趨勢。在實(shí)際生產(chǎn)中為了加快輪蟲繁殖,往往須要設(shè)置高溫,但繁殖速度不代表繁殖能力,溫度越高并不代表種群容納量越大,孫迪杰等探究溫度對褶皺臂尾輪蟲壽命與繁殖的影響,結(jié)果表明,溫度越高繁殖速度越快,但就繁殖能力而言,輪蟲在高溫區(qū)和低溫區(qū)區(qū)別并不顯著[14],本試驗(yàn)結(jié)果與之相符。本試驗(yàn)在以酵母為基礎(chǔ)餌料的情況下,種群容納量在20~30 ℃條件下并非溫度越高越好,原因可能是溫度越接近30 ℃,就越接近2種菌類最適宜的繁殖溫度,在自然生態(tài)環(huán)境下,藻類通過光合作用可產(chǎn)生溶解氧補(bǔ)充水體中的氧氣,供有機(jī)物分解以及水中微生物消耗。而在沒有藻類植物產(chǎn)生光合作用的酵母培養(yǎng)液中,升溫后的枯草芽孢桿菌和酵母菌的快速繁殖會大量消耗氧氣,在沒有采取任何增氧措施的情況下,升高水體溫度后,枯草芽孢桿菌憑借種群繁殖優(yōu)勢再次成為優(yōu)勢種,輪蟲在營養(yǎng)液中唯一的飲食來源被限制,在沒有外界藻類持續(xù)供應(yīng)的情況下,升溫導(dǎo)致水體菌群的改變,從而會對輪蟲的生長和攝食產(chǎn)生影響。總體而言,20、25、30 ℃濃度處理組輪蟲最大密度較對照組分別上升24%、153%、74%,若在生產(chǎn)周期內(nèi)需要在最快速度最短時間內(nèi)得到足量輪蟲,仍然可盡量將溫度控制在高溫(30 ℃)區(qū),若想得到最大生產(chǎn)潛力,則將溫度適當(dāng)降低以獲得最大種群容納量。
4 問題與展望
生物餌料高密度生產(chǎn)過程中,輪蟲從投喂小球藻轉(zhuǎn)移到投喂人工餌料后會大量死亡,其死亡原因與水體中菌落生態(tài)系統(tǒng)突變相關(guān),生物餌料對不同菌類的營養(yǎng)需求以及菌類和主要餌料品種的協(xié)調(diào)作用須要進(jìn)一步探索與研究。本試驗(yàn)從宏觀角度證實(shí)添加枯草芽孢桿菌能明顯提高輪蟲產(chǎn)量,酵母與枯草芽孢桿菌之間的協(xié)同機(jī)制尚不明確,從營養(yǎng)強(qiáng)化角度來分析,長時間食用酵母配合枯草芽孢桿菌能否提高輪蟲體內(nèi)高度不飽和脂肪酸含量還須要進(jìn)一步檢測。其次,輪蟲高密度養(yǎng)殖過程中,水體中菌落生態(tài)較復(fù)雜,單一有益菌種想要形成優(yōu)勢種須要達(dá)到一定數(shù)量,具有一定的難度,所以如何在大水體環(huán)境下確保濃度與活菌數(shù)是生產(chǎn)中的關(guān)鍵,同時對于菌類繁殖環(huán)境的控制也是重點(diǎn),須要盡可能地營造適合菌種的生長環(huán)境,保持養(yǎng)殖水體內(nèi)溶氧環(huán)境,定向培育優(yōu)勢菌群并發(fā)揮其作用,以達(dá)到預(yù)期的效果。如今在高密度養(yǎng)殖導(dǎo)致水體環(huán)境日益惡化的背景下,水產(chǎn)科研人員須要開發(fā)出更多益生菌和微生態(tài)制劑,向水產(chǎn)行業(yè)從業(yè)者傳播健康的微生態(tài)協(xié)調(diào)理念,擴(kuò)大經(jīng)濟(jì)效益和環(huán)保效益,促進(jìn)我國水產(chǎn)育苗行業(yè)的健康發(fā)展。
參考文獻(xiàn):
[1]Hirata H,Mori Y. Cultivation of the rotifer Brachionus plicatilis fed on a mized diet of marine Chlorella and baker s yeast[J]. Saibai Gyogyo,1967,5:36-40.
[2]Hirayama K,Watanabe K. Fundamental studies on physiolgy of rotifer for its mass culture:4. Nutritional effect of yeast on population growth of rotifer[J]. Bull Jap Soc Sci Fish,1973,39(11):1129-1133.
[3]賀詩水. 不同營養(yǎng)強(qiáng)化對褶皺臂尾輪蟲脂類和脂肪酸組成的影響[J]. 糧食與飼料工業(yè),2010(11):58-60.
[4]Satuito C G. Nutritional quality of two species of mirane yeast
(Candia sp. & Saccharomyces sp.) on the population growth of the rotifer Brachionus plicatilis[C]. The Second Asian Fisheries Forum,Manila,Philippines,1990:353-356.
[5]李 磊,朱小明,方 芳,等. 幾種單胞藻對褶皺臂尾輪蟲的營養(yǎng)強(qiáng)化研究[J]. 水產(chǎn)科學(xué),2012,31(2):225-228.
[6]Hirayama,K .Maruyama,Maeda. Nutritional effect of freshwater Chlorella on growth of the rotifer Brachionus plicatilis[J]. Hydrobiology,1989(186/187):39-42.
[7]王金秋,李德尚. 酵母及藻類搭配對萼花臂尾輪蟲餌料效果的研究[J]. 應(yīng)用生態(tài)學(xué)報,1998,9(3):301-304.
[8]王金秋,李德尚,董雙林,等. 面包酵母添加光合細(xì)菌和維生素C培養(yǎng)輪蟲的效果[J]. 水產(chǎn)科技情報,1999,26(1):25-27.
[9]甘松永,黃錦雄,張 敏,等. 利用酵母和芽孢桿菌培養(yǎng)海水輪蟲的技術(shù)初探[J]. 當(dāng)代水產(chǎn),2017,42(11):98-99.
[10]鄧 平,張生元,艾桃山,等. 食物濃度和溫度對萼花臂尾輪蟲種群動態(tài)的影響[J]. 江蘇農(nóng)業(yè)科學(xué),2016,44(1):251-253.
[11]王愛民,席貽龍,牛翔翔,等. 汀棠湖冬季出現(xiàn)的萼花臂尾輪蟲對水溫的適應(yīng)[J]. 生態(tài)學(xué)報,2014,34(16):88-95.
[12]Anna K,Aleksandra W,Edyta F,et al. Seasonal changes in the body size of two rotifer species living in activated sludge follow the temperature-size rule[J]. Ecology & Evolution,2014,4(24):78-89.
[13]鄭樂云. 溫度對超小型輪蟲生長、繁殖及營養(yǎng)成分的影響[J]. 福建水產(chǎn),2010(3):5-9.
[14]孫迪杰,劉娟然. 溫度對褶皺臂尾輪蟲壽命和繁殖的影響[J]. 水產(chǎn)科學(xué),1993,12(6):14-17.肖金星,王 磊,鄭 剛,等. 硒化多糖對黑鯛存活、免疫力和抗氧化性能的影響[J].

- 江蘇農(nóng)業(yè)科學(xué)的其它文章
- 江西省農(nóng)業(yè)產(chǎn)業(yè)集群的區(qū)位商計算與檢驗(yàn)
- 絲綢之路經(jīng)濟(jì)帶核心區(qū)城市化水平與生態(tài)環(huán)境耦合協(xié)調(diào)關(guān)系綜合測度
- 基于因子分析的江蘇省農(nóng)機(jī)政策性保險發(fā)展的區(qū)域差異
- 多維度社會資本視角下農(nóng)戶借貸需求滿足度及其影響因素研究
- 農(nóng)產(chǎn)品圖文呈現(xiàn)與在線評論不一致性對銷量的影響
- 一株產(chǎn)纖維素酶細(xì)菌的發(fā)酵產(chǎn)酶條件優(yōu)化