利用Pdl1 敲除的巨噬細胞轉錄譜分析結核感染中PD-L1 作用相關的候選基因
2020-06-24 08:15:16石亞男李軍麗占玲俊
中國比較醫學雜志 2020年5期
石亞男,唐 軍,李軍麗,王 杰,占玲俊
(中國醫學科學院醫學實驗動物研究所;衛健委人類疾病比較醫學重點實驗室;新發再發傳染病動物模型研究北京市重點實驗室;北京市人類重大疾病實驗動物模型工程技術研究中心;中國醫學科學院結核病中心,北京 100021)
結核病是由結核分枝桿菌(簡稱結核菌,Mycobacterium tuberculosis,Mtb)感染引起的慢性傳染性疾病,是全球十大死因之一。 2019 年世界衛生組織(WHO)報告指出,2017 年全球結核病人數約為17 億,其中新發結核病例1010 萬人,結核病死亡人數約為157 萬[1],WHO 呼吁2030 年終止結核,因此結核的防控研究尤為任重而急迫。
程序性死亡配體1(PD-L1)高表達與活動性結核緊密相關,PD-L1 表達于多種免疫細胞,如巨噬細胞、中性粒細胞、T 細胞和B 細胞[2]。 其中巨噬細胞是結核病肉芽腫形成的主要細胞[3],巨噬細胞作為效應細胞和抗原提呈細胞,在感染過程中通過吞噬殺傷、抗原提呈和分泌多種細胞因子等功能來調控機體炎癥反應和免疫應答[4],是宿主控制Mtb 感染擴散的重要防御屏障。 體外Pdl1 敲低實驗表明,PD-L1 可以通過PD-1: PD-L1/PD-L2 通路在結核感染中發揮免疫抑制作用[5-7],而體內作用機制不明,且體外作用的具體分子機制不明。 除了PD1:PD-L1 通路外,結核病中PD-L1 是否存在類似其他感染或腫瘤疾病中的其它配體和通路,目前還未見報道,其分子機制需要深入研究。
我們通過Cre-loxp 技術構建巨噬細胞特異性Pdl1 敲除小鼠[8],主要目的是在結核感染中,動物體內研究PD-L1 對巨噬細胞的作用機制。 本研究通過比較不同敲除水平Pdl1 的巨噬細胞感染Mtb后轉錄譜變化,從生物信息學篩選到的差異表達基因角度,分析結核菌感染中PD-L1 發揮作用的分子機制,為深入研究PD-L1 在結核感染發病中免疫作用和機制奠定基礎。
1 材料和方法
1.1 實驗材料
1.1.1 實驗菌株
結核分枝桿菌標準株H37Rv(菌號為93009)為本室保存,使用前活化并將菌懸液濃度調整為1.0×107CFU/mL,-80℃保存備用。
1.1.2 實驗動物
3 只4 ~6 周齡SPF 級雄性Pdl1Flox/-小鼠體重12~14 g,由中國醫學科學院醫學實驗動物研究所基因工程平臺采用CRISPR/Cas9 技術構建。 4 ~6周齡SPF 級野生型C57BL/6 小鼠購自北京維通利華[SCXK(京)2017-001],體重12 ~14 g,4 ~6 周齡SPF 級雄性Lyz2-iCre 小鼠由南京大學模式動物中心惠贈,體重12 ~14 g,均飼養于中國醫學科學院醫學實驗動物研究所屏障環境動物房[SYXK(京)2014-0029],動物實驗中涉及動物的操作程序已經得到本所實驗動物使用與管理委員會(IACUC)的批準,批準號為ZLJ19003,所有動物按3R 原則予以關懷。
1.2 主要試劑
中性羅氏培養管(珠海貝索生物,中國);FBS、DMEM 培養基、RPMI-1640 培養基(Gibco,美國);TRIzol Reagent(Life Technologies,美國);DNA 提取試劑盒(EE101-02)購自北京全式金;PCR 引物由上海英濰捷基合成;1×RBC Lysis Buffer、Anti-mouse CD3e FITC(eBioscience,美國);PE/Cy7 anti-mouse CD19、APC anti-mouse CD4、FITC anti-mouse/human CD11b、APC anti-mouse F4/80、PE anti-mouse CD274(B7-H1,PD-L1)(Biolegend,美國)。
1.3 實驗方法
1.3.1 巨噬細胞特異性Pdl1 敲除小鼠制備和繁育
巨噬細胞特異性Pdl1 敲除小鼠的設計利用了位點特異性重組酶系統Cre/loxp 原理,需要在Pdl1基因序列的兩端各放置一個loxp 序列,得到Pdl1Flox/-小鼠。 將Pdl1Flox/-小鼠與巨噬細胞特異的Cre 小鼠交配繁殖,以獲得在巨噬細胞內把Pdl1 基因敲除掉的小鼠,即巨噬細胞特異性Pdl1 敲除小鼠,具體方法如下:
Pdl1Flox/-小鼠構建成功后,與C57BL/6 小鼠進行雜交,得到的Pdl1Flox/-小鼠進行自交,進一步得到Pdl1Flox/Flox小鼠,將雌性Pdl1Flox/Flox小鼠和帶有巨噬細胞特異性表達Cre 酶的Lyz2-iCre 雄鼠進行雜交,得到Pdl1Flox/--Cre 小鼠,最后將Pdl1Flox/Flox小鼠和Pdl1Flox/-- Cre 小 鼠 雜 交, 最 終 得 到 1/4 的Pdl1Flox/Flox- Cre 小鼠,即為巨噬細胞特異性Pdl1 敲除小鼠,同時獲得1/4 的Pdl1Flox/--Cre 小鼠及1/2的同窩陰性Pdl1Flox/Flox小鼠。
1.3.2 小鼠基因型鑒定
剪腳趾標記出生幼鼠,收集腳趾并進行全基因組DNA 提取。 每管加入100 μL LB2 和20 μL 蛋白酶K,56℃金屬浴孵育至完全裂解后參照DNA 提取試劑盒說明書進行,純化后的全基因組DNA 儲存于-20℃冰箱備用。 根據待鑒定樣品數量配制所需的PCR 反應體系,同時設置陰性對照和陽性對照進行PCR 擴增。 其中Pdl1-loxp 上游引物序列為: 5’TGTAAACTATTAGACTGGCTGGAATGTAG 3’;下游引 物 序 列 為: 5’ GATGCAAGCTAGATAATATATT CCCAGTC 3’,目標片段大小為KI 1700 bp。 Cre 上游引物序列為:5’AGTGCTGAAGTCCATAGATCGG 3’;下游引物序列為5’CTGATTCTCCTCATCACC AGG 3’。 目標片段大小為543 bp。 待擴增完畢,分別進行2%和1% 瓊脂糖凝膠電泳鑒定。
1.3.3 PD-L1 表達水平的檢測
(1) 腹腔巨噬細胞PD-L1 表達水平檢測:Pdl1Flox/Flox- Cre、Pdl1Flox/-- Cre、Pdl1Flox/Flox小鼠腹腔注射6%可溶性淀粉肉湯1 mL,3 d 后小鼠斷頸處死,無菌分離腹腔巨噬細胞;1300 r/min、室溫離心10 min,RPMI-1640 培養基洗滌巨噬細胞兩次,1 mL PBS 重懸細胞,分別加入APC-F4/80、FITCCD11b 和PE-CD274 流式抗體,以同型對照PE-IgG為陰性對照,室溫避光孵育15 min,1500 r/min,離心5 min,棄上清;2 mL PBS 洗滌細胞,1 mL PBS 重懸后進行流式細胞術分析以檢測巨噬細胞PD-L1表達水平。
(2)T 細胞與B 細胞PD-L1 表達水平檢測:無菌解剖Pdl1Flox/Flox- Cre、Pdl1Flox/-- Cre、Pdl1Flox/Flox小鼠,脾研磨液過70 μm 孔徑篩網,1500 r/min,離心5 min,棄上清;每管加入5 mL 1×RBC Lysis Buffer,渦旋30 s,室溫靜置4 min;1500 r/min,4℃,離心5 min,棄上清,5 mL PBS 洗滌細胞3 次,1 mL PBS 重懸細胞,分別加入FITC-CD3e、APC-CD4、PE/Cy7-CD19 和PE-CD274 流式抗體,以同型對照PE-IgG為陰性對照,室溫避光孵育15 min,同上進行流式細胞術分析以檢測T 細胞與B 細胞PD-L1 表達水平。
1.3.4 感染與檢測
取24 孔細胞培養板,每孔加入500 μL 新分離的Pdl1Flox/Flox- Cre、Pdl1Flox/-- Cre、Pdl1Flox/Flox小鼠腹腔巨噬細胞,濃度為1×106細胞/mL。 37℃、5%CO2貼壁培養2 d;棄上清,每孔補加500 μL DMEM完全培養基,按MOI 10 加入Mtb 進行感染,未感染對照組每孔加入等體積的DMEM 完全培養基;感染24 h 后,棄上清,1 mL PBS 洗滌貼壁細胞3 次,每孔加入1 mL TRIzol 并收集各組細胞裂解液,提取全基因組進行測序和數據分析。
1.4 統計學方法
基因組比對采用STAR(v2.5.2b)軟件,差異表達分析采用DEGSeq(v1.12.0)軟件,基因本體論(Gene Ontology,GO)富集分析采用GOSeq 軟件(v1.22),京都基因和KEGG 富集分析采用KOBAS(v2.0)軟件。
2 結果
2.1 巨噬細胞特異性Pdl1 敲除小鼠基因鑒定
繁育實驗所需巨噬細胞特異性Pdl1 敲除小鼠,剪新出生小鼠腳趾,提取鼠趾基因組,用PCR 技術進行小鼠基因鑒定。 Pdl1Flox/Flox-Cre 小鼠(圖中標號為7、10 小鼠)及Pdl1Flox/--Cre 小鼠(圖中標號為3、4、6、8、12 小 鼠) 用 于 進 一 步 研 究,同 窩 陰 性Pdl1Flox/Flox小鼠(圖中標號為9 小鼠)作為對照組,圖中標號1、11 為野生型小鼠,2、5 為Pdl1Flox/-小鼠(圖1)。
2.2 PD-L1 表達的檢測結果
流式細胞儀檢測發現:Pdl1Flox/Flox- Cre 小鼠腹腔巨噬細胞PD-L1 的表達量是顯著偏低的,P <0.001(圖2A - 1);三種亞品系Pdl1Flox/Flox-Cre、Pdl1Flox/--Cre、Pdl1Flox/Flox小鼠T 細胞與B 細胞PDL1 的表達量是大致相同的(圖2B-1 與圖2C-1)。因此巨噬細胞特異性Pdl1 敲除小鼠制備成功。
2.3 數據產出質量
測序后Pdl1Flox/Flox- Cre 感染組、Pdl1Flox/-- Cre感染組、Pdl1Flox/Flox感染組及其各自對應的未感染組數據質量通過過濾后的測序數據(Clean reads)大小、99.00%堿基正確識別率(Q20)和99.90%堿基正確識別率(Q30)來判定(表1)。 可見,測序質量能夠滿足后續分析的要求。
2.4 測序序列(reads)與參考基因組比對結果
各組樣品reads 與參考基因組比對的總百分比(Total mapped reads): Pdl1Flox/Flox-Cre 感染組為98.66%,Pdl1Flox/--Cre 感染組為98.70%,Pdl1Flox/Flox感染組為98.57%, 未感染組分別為98.67%,98.69%和98.58%,均高于65.00%,表明參考基因組選擇合適(表2)。
2.5 差異表達分析結果
差異表達結果的分析比較思路是:將每個亞品系的小鼠的感染組與未感染組相比,其差異基因認為是與Mtb 感染相關的候選基因,然后將每個敲除小鼠篩到的與Mtb 感染相關的候選基因與同窩陰性小鼠分別比較,其差異基因再結合Pdl1Flox/Flox-Cre 和Pdl1Flox/--Cre 小鼠共同的Mtb 感染相關基因分析,最終得到結核感染中PD-L1 特異性的基因。
與Pdl1Flox/Flox-Cre 的未感染組相比,Pdl1Flox/Flox-Cre 感染組差異表達的基因共有744 個(Q <0.005),其中上調表達基因415 個,下調表達基因329 個(圖3A);與Pdl1Flox/--Cre 的未感染組比較,Pdl1Flox/--Cre 感染組差異表達的基因共有811 個(Q <0. 005),其中上調表達基因459個,下調表達基因352 個(圖3B);與Pdl1Flox/Flox的未感染組比較,Pdl1Flox/Flox感染組差異表達的基因共有510 個(Q<0.005),其中上調表達基因332 個,下調表達基因178 個(圖3C)。

圖1 巨噬細胞特異性Pdl1 敲除小鼠基因鑒定Note. N, Negative control. P, Positive control. M, DNA marker.Figure 1 Genotype identification of macrophage-specific Pdl1 conditional knockout mice
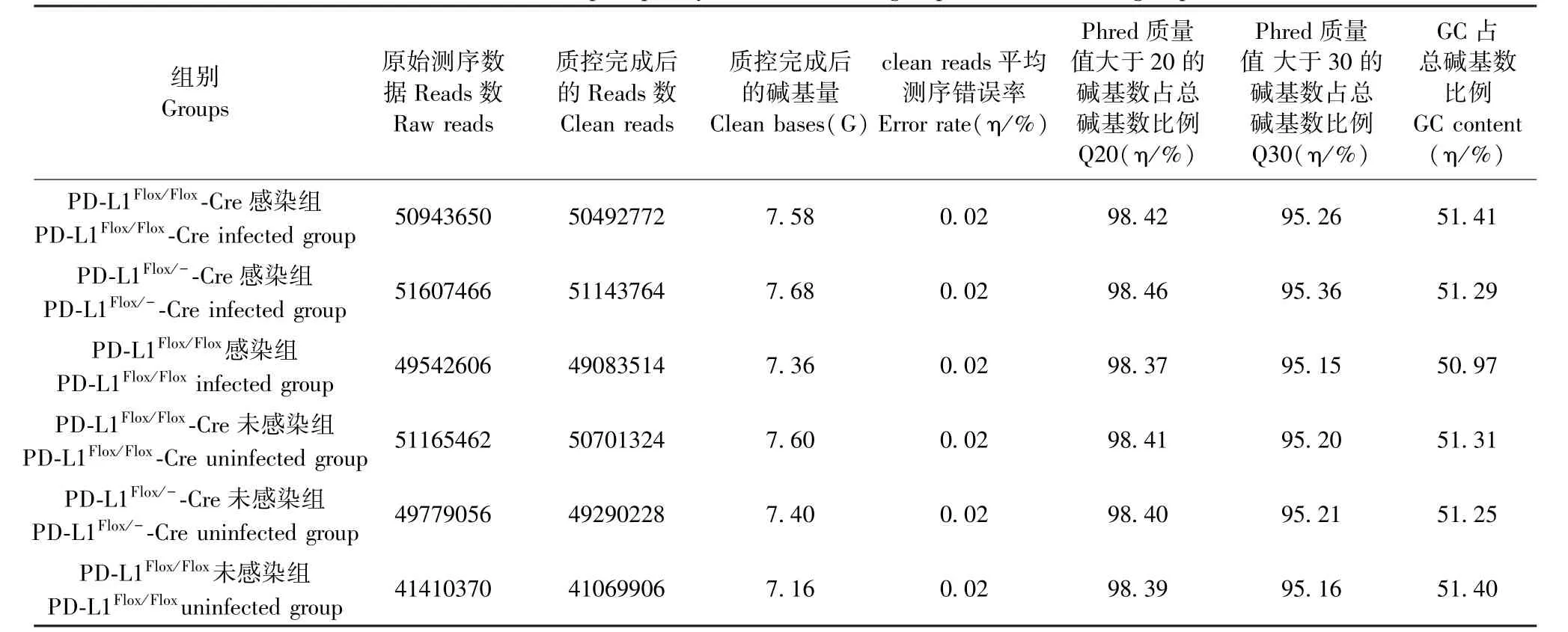
表1 感染組和未感染組數據產出質量Table 1 Data output quality list in infected group and uninfected group
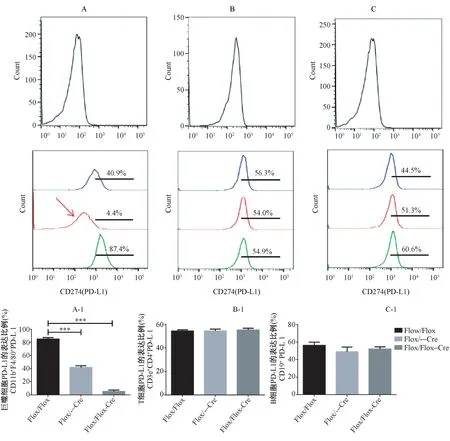
圖2 流式細胞術檢測巨噬細胞、T、B 細胞上PD-L1 的表達Note. A, The above stands for the negative control of macrophages, and the below is the expression of PD-L1 in peritoneal macrophages of three sub-lineage mice. B, The above stands for the negative control of B cells, and the below is the expression of PD-L1 in B cells of three sub-lineage mice. C,The above stands for the negative control of T cells, and the below is the expression of PD-L1 in T cells of three sub-lineage mice. The blue peak represents Pdl1Flox/-with Cre mice, the red peak indicates Pdl1Flox/Floxwith Cre mice, and the green peak represents Pdl1Flox/Flox mice. Compared with the Pdl1Flox/Flox group,***P <0.001.Figure 2 The expression levels of PD-L1 on macrophages, T cells,and B cells were detected by flow cytometry
2.6 差異表達基因的GO 富集分析結果
與未感染組相比,Pdl1Flox/Flox-Cre 感染組,Pdl1Flox/--Cre 感染組,Pdl1Flox/Flox感染組最顯著富集組份均為參與免疫反應的GO:0006955 (P <0. 01)與參與免疫系統過程的GO:0002376(P <0. 01),涉及差異表達的基因分別為18 個(表3),28 個(表4),20 個(表5),其中Pdl1Flox/Flox-Cre 感染組與Pdl1Flox/--Cre 感染組共有32 個差異表達基因,其中30 個基因表達上調,2 個基因表達下調。 這些共有的差異表達基因進一步與Pdl1Flox/Flox感染組進行比較,剔除相同基因,篩選出的候選基因初步定為結核感染中PD-L1 相關的特異基因,共17 個基因,其中13 個基因表達上調,4 個基因表達下調。
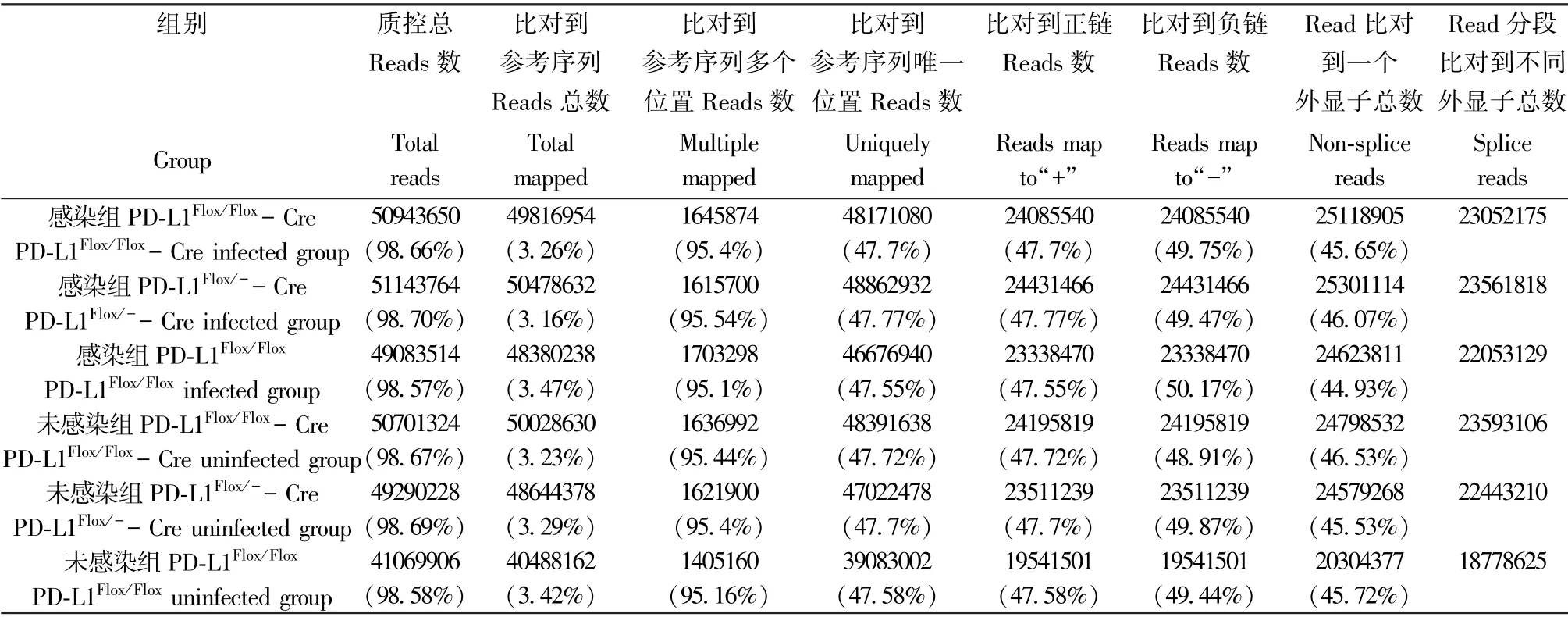
表2 感染組和未感染組reads 與參考基因組比對情況Table 2 Comparison of reads and reference genomes between infected group and uninfected group

圖3 差異基因火山圖Note, A, The Pdl1Flox/Flox with Cre infected group compared with the uninfected group. B, The Pdl1Flox/- with Cre infected group compared with the uninfected group. C, The Pdl1Flox/Floxinfected group compared with the uninfected group. The abscissa represents the fold change in gene expression in different samples,and the ordinate represents the statistically significant differences in gene expression.The significantly differentially expressed genes are represented by red dots (upregulation) and green dots (downregulation). Genes that were not significantly differentially expressed are indicated by blue dots.Figure 3 Differential gene volcano map
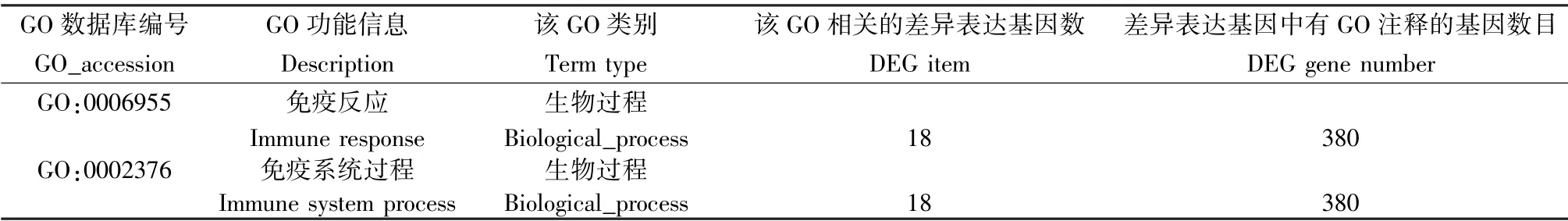
表3 PD-L1Flox/Flox-Cre 感染組與其未感染組相比差異表達基因GO 最顯著富集分析的前2 項Table 3 Most two enriched terms of GO enrichment analysis of differentially expressed genes in PD-L1Flox/Flox-Cre infected group compared with its uninfected group
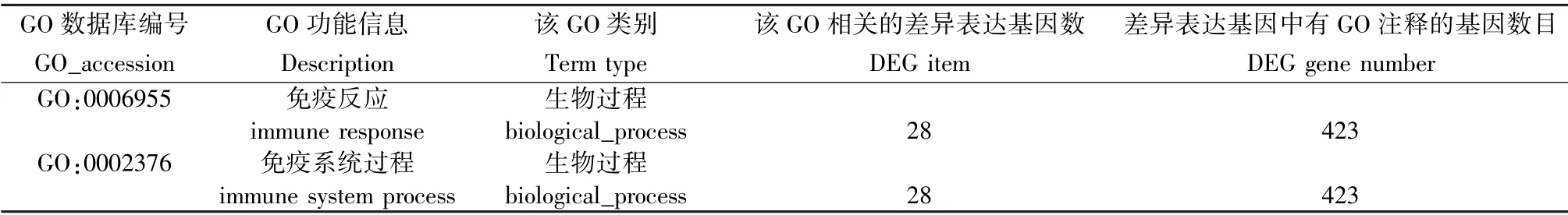
表4 PD-L1Flox/--Cre 感染組與其未感染組相比差異表達基因GO 最顯著富集分析的前2 項Table 4 Most two enriched terms of GO enrichment analysis of differentially expressed genes in PD-L1Flox/--Cre infected group compared with its uninfected group
上調和下調表達基因名稱及對應的變化倍數對數值[|log2(FoldChange)|]整理如下:上調表達的基因包括促炎因子趨化因子配體2(Ccl2,3.4677)、趨化因子配體7(Ccl7,8.1765)、白介素12a(Il12a,6.1789)、白介素6(Il6,9.0296)、趨化因子(C-X-C 模體)配體1(Cxcl1,6.1104)、趨化因子(C-X-C 模體)配體9(Cxcl9,1.9734)、組織相容性2,Q 區基因座5(Qa5,1.3648)、組織相容性2,M 區基因座2(Hmt,1.2766)、組織相容性2,T 區基因座23(Qa1, 1.1328)、 鳥苷酸結合蛋白4 (Gbp4,4.6111)、鳥苷酸結合蛋白8(Gbp8,2.8399)、鳥苷酸結合蛋白9(Gbp9,4.9383)、鳥苷酸結合蛋白11(Gbp11,6.649)。 下調表達的基因包括趨化因子(C-C 模體)配體9(Ccl9,1.3723)、白介素1 受體拮抗劑(Il1rn,1.9407)、補體成分1q 亞成分,α 多肽(C1qa,1.392) 和組織相容性2,K 區基因座2(H2k2,1.2708)。 上述基因表明Mtb 感染Pdl1 敲除的巨噬細胞24 h 后,可能通過上述免疫分子參與結核的特異性免疫反應。
2.7 差異表達基因的KEGG 富集分析結果
通過京都基因和基因組百科全書(Kyoto encyclopedia of Genes and Genomes,KEGG)富集分析,發現與未感染組相比,Pdl1Flox/Flox-Cre 感染組最顯著富集的通路為Toll 樣受體(Toll-like receptors)信號通路、核因子κB(nuclear factor kappa-B,NFκB)信號通路及缺氧誘導因子-1(HIF-1)信號通路(P<0.01,表6);Pdl1Flox/--Cre 感染組最顯著富集的通路為腫瘤壞死因子(TNF)信號通路及NF-κB 信號通路(P<0.01,表7);Pdl1Flox/Flox感染組最顯著富集的通路為TNF 信號通路及NF-κB 信號通路(P<0.01, 表 8)。 其 中 Pdl1Flox/Flox-Cre 感 染 組 與Pdl1Flox/--Cre 感染組富集的通路中共有16 個差異表達基因,表達均上調,這些共有的差異表達基因進一步與Pdl1Flox/Flox感染組進行比較,剔除相同基因,篩選出6 個基因,表達均上調,基因名稱及變化倍數對數值[| log2(FoldChange)|]整理如下:CD40 抗原(Cd40,3.9838)、TNF 受體相關因子1(Traf1,2.2187)、 TNF 受體相關因子2 (Traf2,1.3572)、TNF 受體相關因子3(Traf3,1.3196)、前列腺素-內過氧化物合酶2(Ptgs2,7.7714)、Caspase募集結構域11(Card11,3.3684),上述基因初步定為結核感染中PD-L1 相關的特異基因。
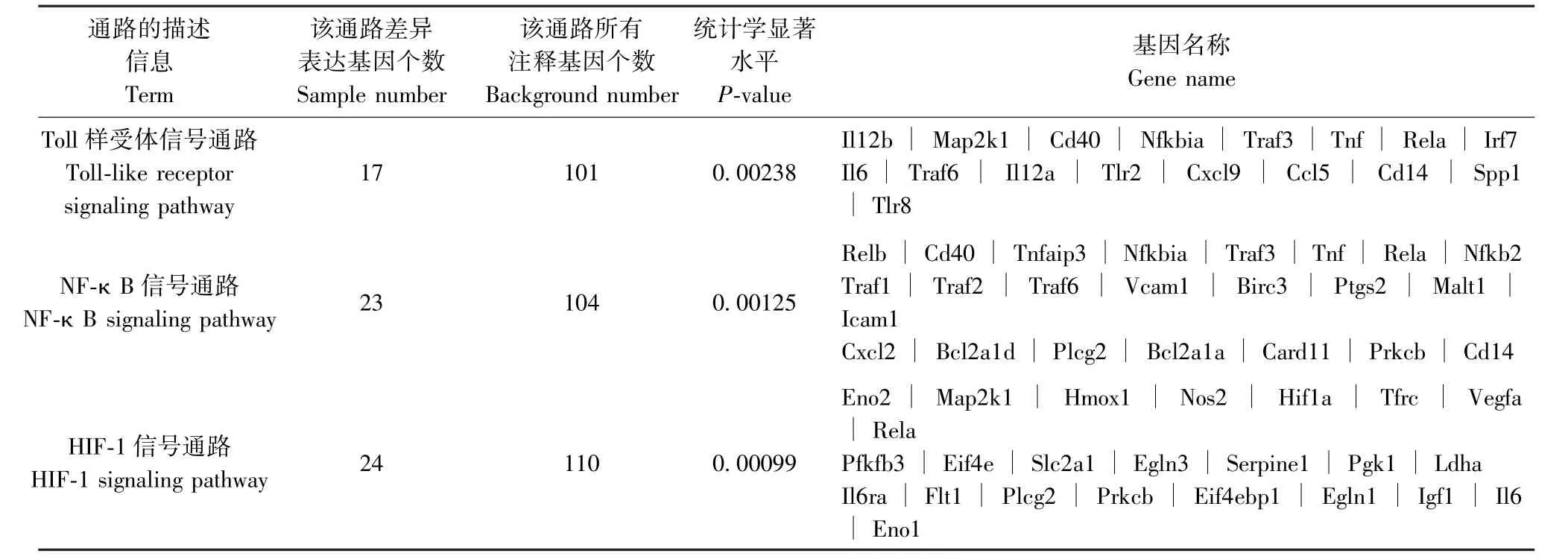
表6 PD-L1Flox/Flox-Cre 感染組與其未感染組相比差異表達基因KEGG 最顯著富集分析Table 6 Most enriched pathway terms of KEGG enrichment analysis of differentially expressed genes in PD-L1Flox/Flox-Cre infected group compared with its uninfected group
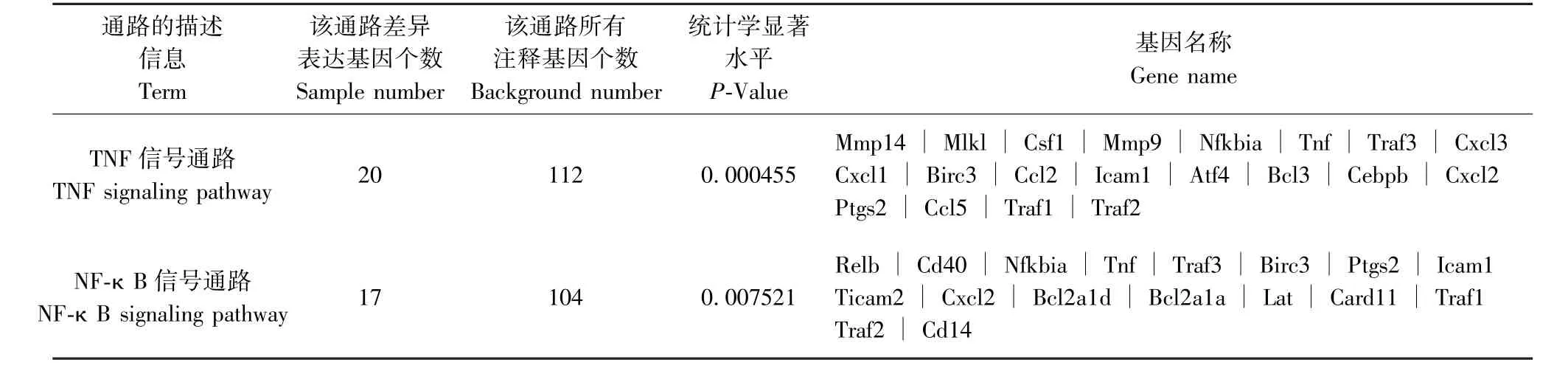
表7 PD-L1Flox/--Cre 感染組與其未感染組相比差異表達基因KEGG 最顯著富集分析Table 7 Most enriched pathway terms of KEGG enrichment analysis of differentially expressed genes in PD-L1Flox/--Cre infected group compared with its uninfected group
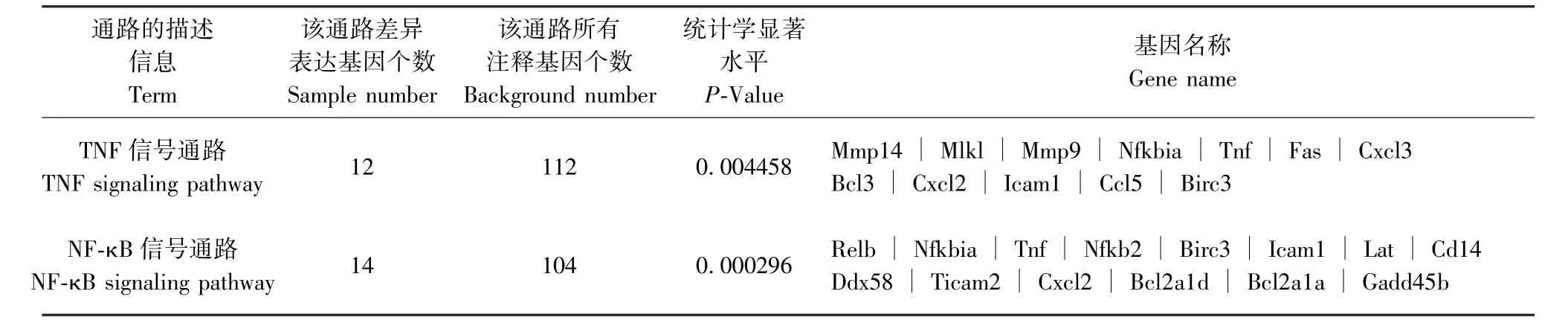
表8 PD-L1Flox/Flox感染組與其未感染組相比差異表達基因KEGG 最顯著富集分析Table 8 Most enriched pathway terms of KEGG enrichment analysis of differentially expressed genesin PD-L1Flox/Flox infected group compared with its uninfected group
3 討論
巨噬細胞在宿主抗結核分枝桿菌的感染免疫中起關鍵作用[9],Mtb 通過抑制吞噬體成熟、抑制吞噬體和溶酶體融合、抑制巨噬細胞凋亡、干擾抗原呈遞、抑制自噬等,逃逸巨噬細胞對Mtb 的殺傷[10],但其具體機制和過程尚未闡明。 研究發現在活動性結核病人中,PD-L1 在中性粒細胞(PMN)和單核/巨噬細胞(Mo/MΦ)表達升高最顯著,PD-L1 表達下降與活動性結核的好轉有密切關系[11],體外實驗發現:PD-1/PD-L1 通路阻斷后,巨噬細胞對Mtb的吞噬和胞內殺傷活性明顯增強[12],然而具體分子機制不明。 此外,PD-L1 是否有其他受體配體通路,目前尚未證實,因此探討PD-L1 在Mtb 感染的巨噬細胞中的作用對結核防治的基礎研究有重要意義。
本研究通過Cre-loxp 技術構建巨噬細胞特異性Pdl1 敲除小鼠模型,通過PCR 和流式細胞術檢測證實巨噬細胞特異性Pdl1 敲除小鼠構建成功。 分離腹腔巨噬細胞,對Mtb 感染前后的不同敲除小鼠的巨噬細胞進行RNA-Seq 及生物信息學分析,結果表明:Mtb 感染的Pdl1 敲除巨噬細胞中基因表達譜發生改變,通過GO 和KEGG 富集分析,初步篩選出23個PD-L1 相關的候選基因,其中某些基因在腫瘤和病毒感染中已有相關報道,在人卵巢癌和結腸癌中,阻斷PD-1/PD-L1 信號通路,可促進Th1 型趨化因子CXCL9 的分泌[13],促進效應T 細胞和自然殺傷(NK)細胞浸潤,增強抗腫瘤效應。 在神經膠質瘤、黑色素瘤微環境中,CCL2、CXCL1、PTGS2 及CD40 的表達與PD-L1 的表達呈正相關[14-17],阻斷PD-1/PD-L1 信號通路,可促進腫瘤相關巨噬細胞(TAM)釋放促炎細胞因子IL6,增強Th1 免疫應答和抗腫瘤作用[18]。 在人丙型肝炎病毒(HCV)感染中,阻斷PD-1/PD-L1 信號通路,可增強低表達TRAF1 的特異性CD8+T 細胞反應[19]。 如前所述,在結核分枝桿菌感染中,抑制PD-1/PD-L1 信號通路可促進小鼠肺泡巨噬細胞釋放炎性因子IL6,增強先天免疫反應[12]。
通過KEGG 富集分析,發現與PD-L1 有關的通路包括Toll 樣受體信號通路、NF-κB 信號通路、HIF-1 信號通路及TNF 信號通路。 在Mtb 感染初期,Toll樣受體作為天然免疫細胞活化的調節劑發揮著重要作用,隨后,Toll 樣受體及信號通路對特異性細胞免疫產生影響。 此外,Toll 樣受體信號通路亦參與調控結核菌免疫逃逸的發生,是導致結核慢性感染的根源所在[20]。 NF-κB 是一類重要的核轉錄因子,其活化可激活下游眾多的細胞因子的轉錄,NF-κB信號通路和NF-κB 依賴性細胞因子在遏制結核菌感染中至關重要[21]。 HIF-1 是一種重要的轉錄因子,是由HIF-1α 和HIF-1β 組成的異源二聚體,其中HIF-1α 為主要的調節亞基,在缺氧、腫瘤及炎癥情況下可廣泛表達[22],是抗結核分枝桿菌感染的重要介質[23]。 TNF 信號傳導通路主要包括Caspase 家族介導的細胞凋亡、銜接蛋白TRAF 介導的轉錄因子NF-κB 和JNK 蛋白激酶的活化,在抗結核感染中發揮重要作用[24]。
本研究通過GO 富集分析發現的免疫反應有關的17 個候選基因與KEGG 富集分析發現的免疫及炎癥相關代謝通路的6 個候選基因,可能是結核感染中PD-L1 作用相關的特異性分子,后期將對這23個基因進行細胞水平和動物體內的驗證和功能機制研究,為結核防治的基礎研究提供線索。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
音樂探索(2022年2期)2022-05-30 21:01:37
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
