基于線粒體COⅠ基因序列的江蘇省4個太湖新銀魚群體遺傳多樣性分析
2020-06-30 08:08:50李大命唐晟凱劉燕山谷先坤劉小維殷稼雯張彤晴潘建林
海洋漁業 2020年3期
李大命,唐晟凱,劉燕山,谷先坤,劉小維,殷稼雯,王 彬,馬 昊,張彤晴,潘建林
(江蘇省淡水水產研究所,江蘇省內陸水域漁業資源重點實驗室,南京 210097)
太湖新銀魚(Neosalanx taihuensisChen,1956)隸屬于鮭形目(Salmoniformes)銀魚科(Salangidae)新銀魚屬,為1年生小型魚類,具有很高的營養價值,主要分布在我國黃河、淮河和長江中下游及其附屬湖泊,是我國重要的經濟魚類[1]。太湖新銀魚生活周期短、生長快、世代離散,對環境變化敏感,種群波動顯著,是典型的r-對策生物[2]。多年來,受圍湖造田、過度捕撈、環境污染和生境破碎化等多種因素的影響,我國的太湖新銀魚天然資源持續衰退,分布范圍顯著縮小,保護和恢復太湖新銀魚種質資源成為亟待解決的問題[2]。
遺傳多樣性是生物多樣性的核心和重要組成部分,了解和掌握生物的遺傳多樣性是制定物種保護策略的前提。只有在了解種群的遺傳背景和現有的遺傳格局及成因的基礎上,才能為制定物種保護策略提供科學依據[3-4]。線粒體DNA具有結構簡單、母系遺傳、進化速率快、幾乎不發生重組等特點,成為研究種群遺傳多樣性的常用分子標記之一[5]。其中,細胞色素c氧化酶亞基 I(cytochrome oxidase subunit I,COⅠ)序列長度適宜,進化速度較快,密碼子的保守性高且引物的通用性強,其不僅適合于種群水平遺傳多樣性的檢測,也可用于種間分析,被廣泛應用于魚類的分子系統學和種群遺傳學研究[6-8]。
江蘇省位于長江、淮河下游,湖泊眾多,是我國銀魚資源的重要分布區域。太湖、洪澤湖、高郵湖和駱馬湖是江蘇省的四大淡水湖泊。歷史上,四大淡水湖泊銀魚資源豐富,不僅是湖泊漁業生產的重要捕撈對象,也曾是出口創匯的重要水產品,但近年來銀魚資源逐漸趨于枯竭[9]。我國在包括太湖新銀魚在內的銀魚科魚類的形態學、個體生物學及生態學方面已進行了廣泛的研究[1-2],但相關的遺傳學資料較少。已有文獻報道利用線粒體Cytb、COⅠ等分子標記分析我國太湖新銀魚的遺傳多樣性和遺傳結構[10-11]。目前,太湖、洪澤湖的銀魚遺傳多樣性已有報道[12-15],但尚未見有關高郵湖和駱馬湖的銀魚遺傳多樣性研究。本文選用COⅠ基因作為分子標記,利用PCR擴增和測序技術,全面分析和比較江蘇省太湖新銀魚種群的遺傳多樣性水平,探究太湖新銀魚種群的遺傳結構特征。研究結果一方面有利于豐富太湖新銀魚的遺傳學資料,掌握太湖新銀魚遺傳資源現狀;另一方面可以為太湖新銀魚種質資源管理保護和開發利用提供科學依據。
1 材料和方法
1.1 實驗材料
2018年8月至12月利用拖網采集太湖、高郵湖、洪澤湖和駱馬湖的太湖新銀魚野生群體,其中太湖群體38尾樣本,高郵湖群體40尾樣本,洪澤湖群體40尾樣本,駱馬湖群體35尾樣本,共153尾樣本(圖1)。剪取太湖新銀魚肌肉組織,放入1.5 mL離心管中,加入無水乙醇保存,帶回實驗室備用。
1.2 DNA提取、PCR擴增與測序
采用Takara公司的基因組DNA提取試劑盒提取太湖新銀魚基因組DNA,將DNA溶于TE溶液中。采用1%瓊脂糖凝膠電泳檢測所提取DNA的完整性,用核酸蛋白定量儀檢測其濃度。
擴增COⅠ序列的正向引物為:F1(5′-TCAACCAACCACAAAGACATTGGCAC-3′),反 向引物為:R1(5′-TAGACTTCTGGGTGGCCAAAGAA TCA-3′)[16]。PCR擴增體系為50μL,Taq酶混合液25μL,上下游引物各2μL(10μmol),DNA模板(40 ng·μL-1)2μL,用 ddH2O補足至 50μL。PCR擴增條件:94℃預變性3 min;94℃變性30 s,55℃退火 40 s,72℃延伸 50 s,30個循環;72℃最后延伸10 min。
用1.5%的瓊脂糖凝膠檢測PCR產物,凝膠成像系統拍照。PCR產物送往生工生物工程(上海)股份有限公司進行雙向測序,測序采用與PCR反應相同的引物。
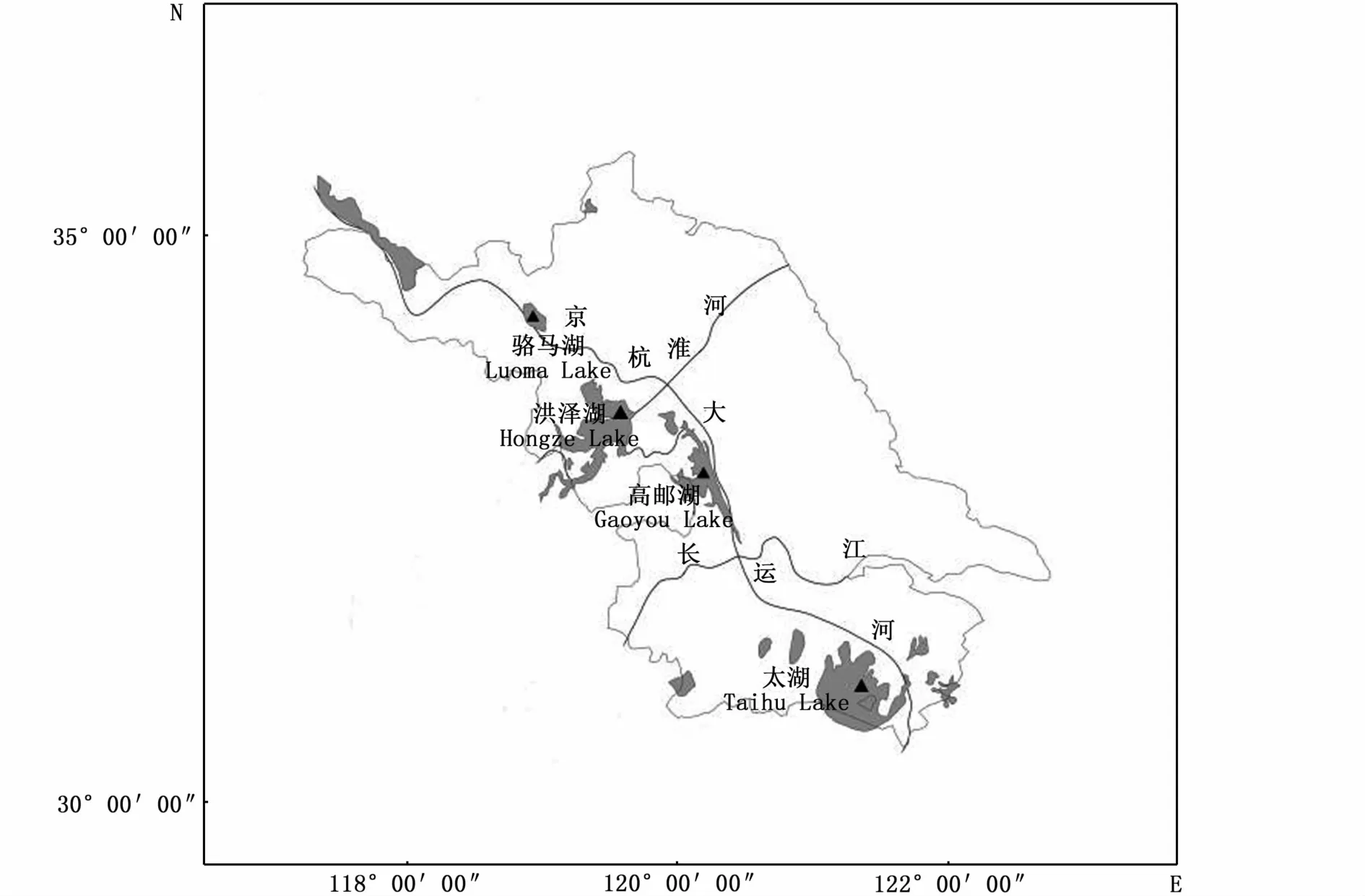
圖1 樣本采集地點Fig.1 M ap of sampling locations
1.3 數據分析
采用BioEdit 7.0軟件[17]對測序結果進行編輯和同源比對。采用 DnaSP 5.0軟件[18]統計核苷酸變異位點、單倍型數目、單倍型多樣性指數和核苷酸多樣性指數。
利用MEGA 7.0軟件[19]統計序列的堿基組成,計算群體間的Kimura雙參數模型(Kimura 2 parameter,K2P)遺傳距離,基于鄰接法(neighborjoinning,NJ)構建單倍型進化樹。采用 Network 4.6.1.0軟件[20]構 建 單 倍 型的 簡 約 中 介(reduced-median,MJ)網絡圖。
使用 Arlequin3.1軟件[121]計算兩兩群體間的遺傳分化指數(Fst),并采用AMOVA來分析群體內及群體間的遺傳結構和分子方差,并通過1 000次重抽樣來檢驗Fst值的顯著性。采用Tajima’sD檢驗[22]和 Fu’sFs檢驗[23]和核苷酸不配對分布(mismatch distribution)分析來檢驗太湖新銀魚群體的歷史動態,以確定是否存在瓶頸效應或群體擴張。
2 結果與分析
2.1 COⅠ基因序列變異和群體遺傳多樣性
本研究共得到153條太湖新銀魚的COⅠ基因序列片段,長度為630 bp。153條COⅠ基因序列共檢測到7個變異位點,其中單一信息位點有4個,簡約信息位點3個,沒有插入或缺失位點。全部序列中堿基A、C、T、G的平均含量分別為19.6%、34.3%、25.3%和 20.8%,其中堿基 A的含量最低,堿基C的含量最高,表現出很強的堿基組成偏向性。
整體來看,太湖新銀魚群體的單倍型多樣性為 0.580±0.022,核苷酸多樣性為 0.001 06±0.000 07(表1)。4個群體中,洪澤湖群體的遺傳多樣性最低,其單倍型多樣性和核苷酸多樣性分別為0.050±0.047和0.000 08±0.000 07;駱馬湖群體的遺傳多樣性最高,其單倍型多樣性和核苷酸多樣性分別為0.361±0.103和0.000 62±0.000 20,但均表現為較低的單倍型多樣性和核苷酸多樣性特點。
2.2 單倍型分布及遺傳關系
4個群體共定義了 9個單倍型(Hap1~Hap9)(表2),其中太湖和洪澤湖群體各擁有2個單倍型,高郵湖群體擁有3個單倍型,駱馬湖群體擁有6個單倍型。9個單倍型中有3個是共享單倍型,其中Hap1被太湖、洪澤湖和駱馬湖群體共享,Hap3和Hap4被高郵湖和駱馬湖群體共享;6個是群體特有單倍型,其中Hap2是洪澤湖群體特有單倍型,Hap5是太湖群體特有單倍型,Hap6是高郵湖群體特有單倍型,Hap7、Hap8和Hap9是駱馬湖群體特有單倍型。

表1 太湖新銀魚遺傳多樣性參數Tab.1 Genetic diversity parameters of four N.taihuensis populations

表2 COⅠ基因單倍型在太湖新銀魚各群體中的分布及數量Tab.2 Distribution and number ofm tDNA COⅠhaplotypes in each population of N.taihuensis
利用Mega 7.0軟件計算9個單倍型之間的Kimura雙參數遺傳距離為0.001 6~0.004 8。采用鄰接法(neighbor-joining,NJ)構建單倍型分子分子系統發育樹(圖2)。從圖2可以看出,NJ進化樹由3個譜系分支構成:單倍型Hap3、Hap6、Hap7、Hap8和 Hap9組成一支(譜系 I),單倍型Hap4和Hap5組成一支(譜系II),單倍型 Hap1和Hap2組成一支(譜系III),但分支支持率較低。4個群體的單倍型相互混雜分布,沒有表現出明顯的地理聚群。
2.3 群體遺傳結構
利用Mega7.0軟件計算4個太湖新銀魚群體間的遺傳距離(表3),結果顯示,4個群體間的遺傳距離均較小,其中太湖群體和洪澤湖群體之間的遺傳距離最小,為0.000 12;太湖群體和高郵湖群體之間的遺傳距離最大,為0.001 86。
AMOVA分子變異方差分析結果顯示(表4),群體間分子變異占76.84%,群體內分子變異占23.16%,分子變異主要發生在群體間。總的遺傳分化系數Fst=0.768 43,且統計檢驗具有極顯著性(P=0.000 0),說明群體間出現了極顯著的遺傳分化。進一步比較兩兩群體間的遺傳分化系數(表3),結果顯示,太湖和洪澤湖群體之間及高郵湖與駱馬湖群體之間的Fst統計檢驗均不顯著(P>0.05),而太湖和洪澤湖群體與高郵湖和駱馬湖群體之間的Fst統計檢驗均極顯著(P<0.001)。
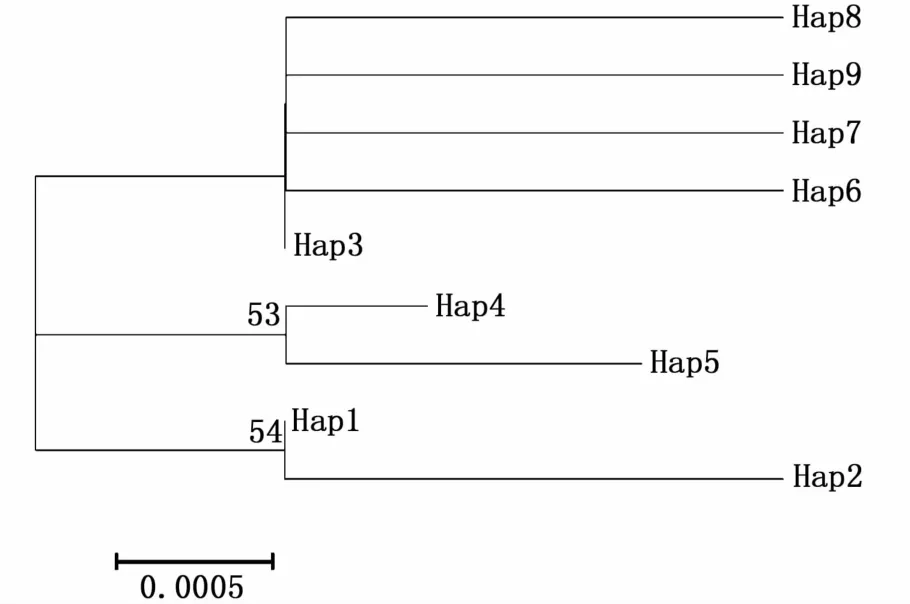
圖2 基于COⅠ基因構建的太湖新銀魚單倍型鄰接樹Fig.2 Neighbor-joining tree of 9 haplotypes from four N.taihuensis populations based on COⅠgene
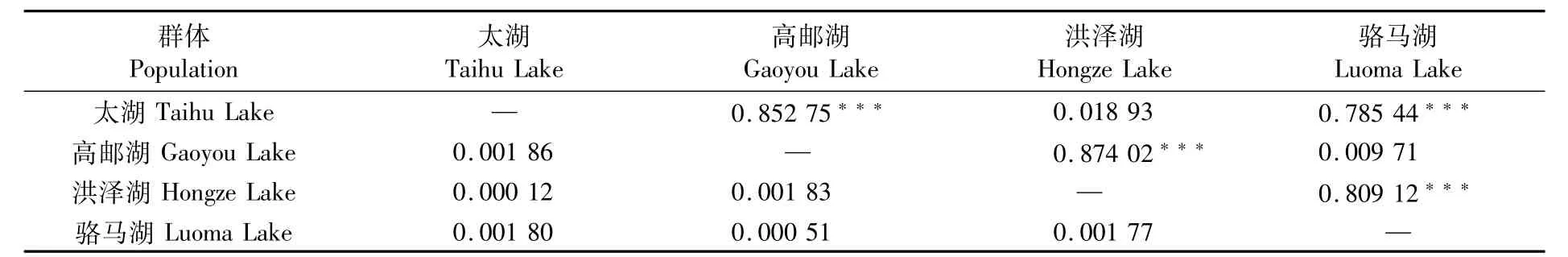
表3 太湖新銀魚群體間遺傳距離(對角線下)和遺傳分化系數(對角線上)Tab.3 Genetic distances(below diagonal)and fixation index(above diagonal)among populations of N.taihuensis

表4 太湖新銀魚群體分子方差分析結果Tab.4 Results ofmolecular variances analysis(AMOVA)of N.taihuensis populations
采用 Network 4.6.1.0軟件構建單倍型之間最小網絡進化關系圖(圖3),結果顯示,太湖和洪澤湖群體的單倍型遺傳關系較近,組成一個進化單元;高郵湖和駱馬湖群體的單倍型遺傳關系較近,組成一個進化單元。單倍型最小網絡進化關系圖進一步支持了分子變異方差分析結果。
2.4 群體歷史動態
對4個太湖新銀魚群體進行中性檢驗(表5)和歧點分布圖分析(圖4),結果顯示,中性檢測Fu’sFs及Tajima’sD的值均為負值,且洪澤湖和駱馬湖群體的檢驗統計結果具有顯著性(P<0.05);另外,4個群體的歧點分布圖呈單峰形,表明4個太湖新銀魚群體進化過程偏離中性,經歷了種群擴張。
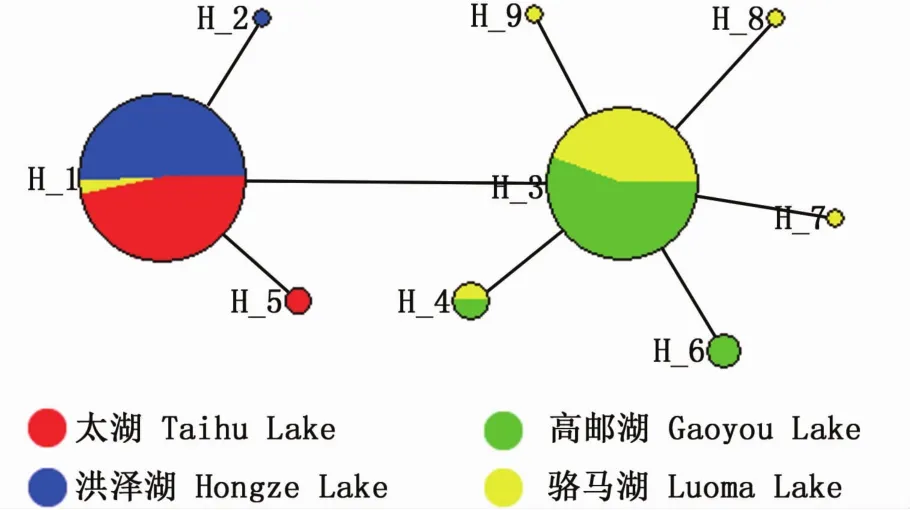
圖3 太湖新銀魚單倍型最小網絡進化關系圖Fig.3 Them inimum spanning network for N.taihuensis hap lotypes
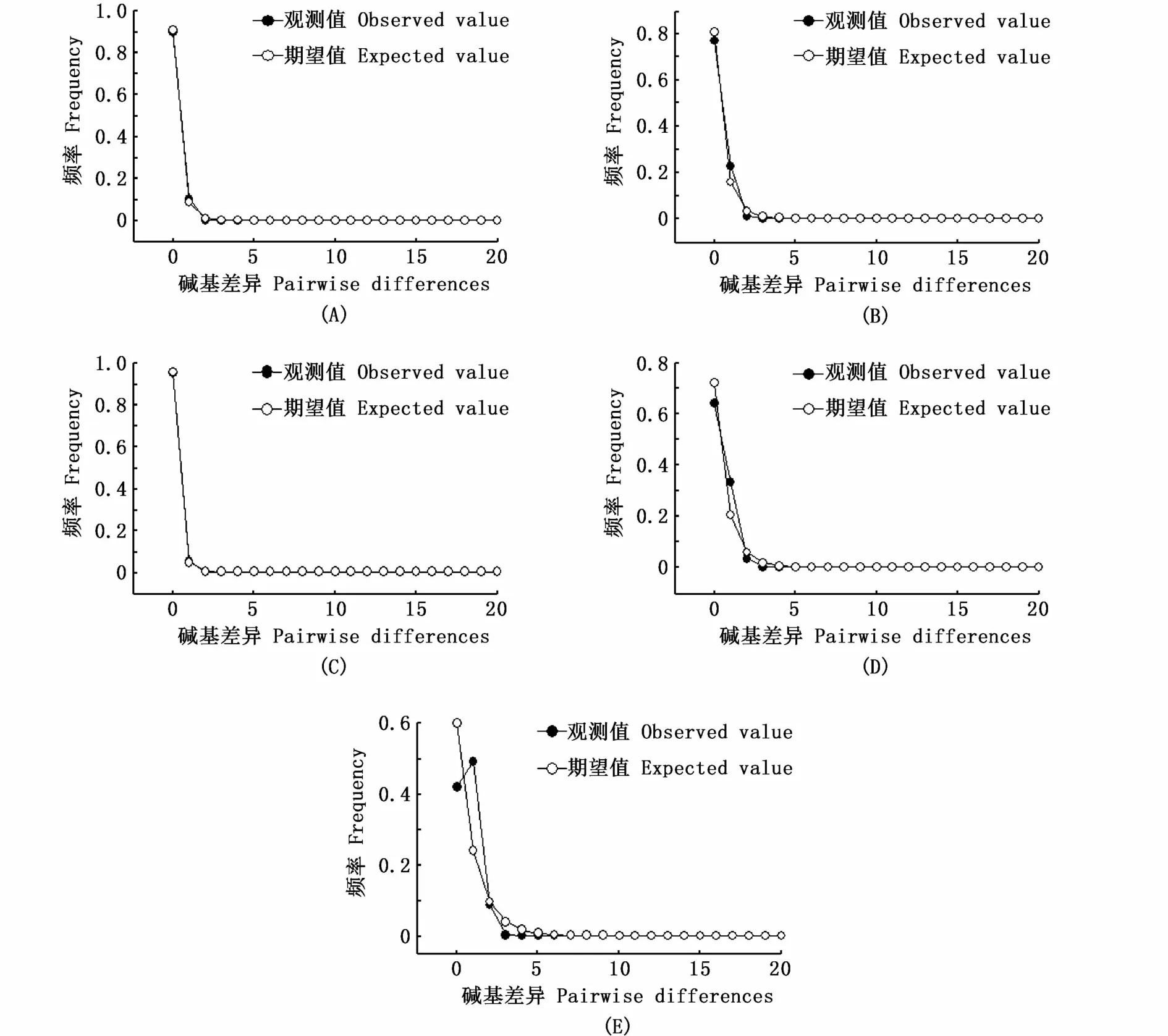
圖4 太湖新銀魚歧點分布圖Fig.4 M ismatch distribution of four N.taihuensis populations
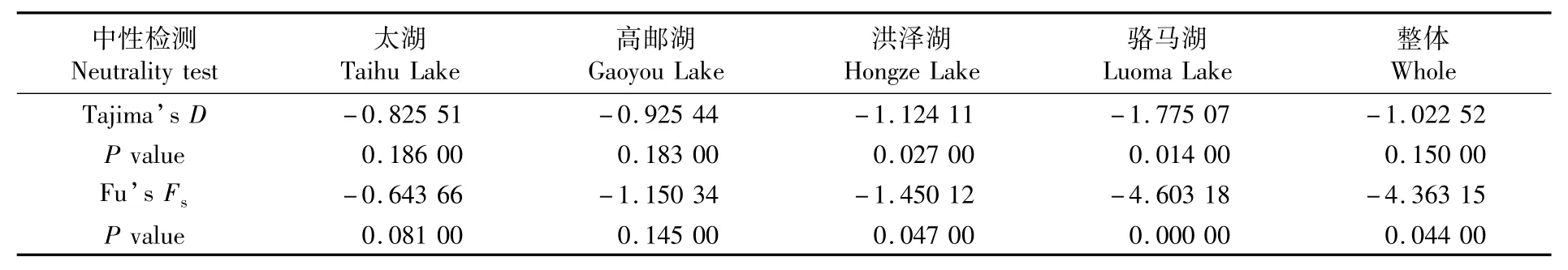
表5 基于COⅠ基因的太湖新銀魚種群中性檢驗Tab.5 Neutrality tests of N.taihuensis populations based on COⅠgene
3 討論
3.1 太湖新銀魚種群的遺傳多樣性
物種遺傳多樣性水平的高低與其適應能力、生存能力和進化潛力存在密切的相關性,遺傳多樣性的降低必將給物種種質資源保護和利用帶來不利影響[24]。通常認為單倍型多樣性指數(h)和核苷酸多樣性指數(π)是衡量一個物種群體多樣性水平的兩個重要指標,與種群大小、年齡結構、近緣物種基因漸滲、群體的突變和選擇有著密切的關系[25]。本研究結果顯示,基于COⅠ序列的4個太湖新銀魚群體的單倍型多樣性為0.050±0.047~0.361±0.103,核 苷 酸 多 樣 性 為0.000 08±0.000 07~0.000 62±0.000 20(表1)。GRANT和 BOWEN[26]提出單倍型多樣性和核苷酸多樣性大小的標準,其中單倍型多樣性以0.5為臨界值,核苷酸多樣性以0.005為臨界值,兩個值越大,物種的遺傳多樣性就越高。比較發現,江蘇省4個太湖新銀魚群體的單倍型多樣性均小于0.5,核苷酸多樣性均小于0.005,表明太湖新銀魚群體的遺傳多樣性處于較低水平。近幾十年來,由于過度捕撈、環境污染、棲息地破壞等不利因素的影響,太湖新銀魚天然資源急劇衰減,種群數量明顯減少,個體組成向小型化、低齡化發展[2,9],導致了太湖新銀魚種群遺傳多樣性喪失。本實驗室對江蘇省五大淡水湖泊漁業資源開展了近十年的連續監測,結果也顯示太湖新銀魚資源量不斷減少,目前已處于極度匱乏狀態①實驗室內部資料,未公開發表數據。。另有文獻報道了基于COⅠ基因的長江中下游太湖新銀魚不同地理群體的遺傳多樣性水平,結果表明,除巢湖 (h:0.667±0.113,π:0.001 60±0.001 91)群體外,洞庭湖(h:0.125±0.106,π:0.000 40±0.000 96)、梁子湖(h:0,π:0)、鄱陽湖(h:0.468±0.101,π:0.000 81±0.001 22)、泊湖(h:0.233±0.126,π:0.000 37±0.000 48)及南漪湖 (h:0.458±0.095,π:0.000 73±0.000 48)群體的遺傳多樣性均呈現出較低水平,本研究結果與之相似[11]。同時,與基于COⅠ基因的太湖大銀魚 (Protosalanx chinensis)(h:0.577,π:0.001 05)和 洪 澤 湖 大 銀 魚 (P.hyalocronius)(h:0.660,π:0.001 54)[14]及基于Cytb基因的喬什新銀魚(N.jordani)(h:0.590±0.047,π:0.000 88±0.000 11)[12]遺傳多樣性相比,太湖新銀魚的遺傳多樣性較低。由此可以推測,我國太湖新銀魚遺傳多樣性較低是一個普遍現象,應引起足夠重視。
3.2 太湖新銀魚種群的歷史動態
一般來說,物種的遺傳多樣性模式與其進化歷史密切相關。當h大于或等于0.5、π小于0.005時,是受瓶頸效應后種群數量的迅速擴張導致;當h大于等于0.5、π大于或等于0.005時,表示種群穩定,具有比較悠久的進化歷史;當h小于0.5、π大于或等于0.005時,種群經歷了輕微的瓶頸效應,幾乎沒有影響到核苷酸變異;當h小于0.5、π小于0.005時,表明種群近期經歷了瓶頸效應[26]。本研究中的4個湖泊群體單倍型多樣性均小于0.5,核苷酸多樣性均小于0.005,表明太湖新銀魚最近可能經歷了嚴重的瓶頸效應。
種群歷史演化通常通過兩種方法來檢測:其一是堿基不配對分析,該方法是基于無限位點模式(infinite-sitemodel)對樣本中兩兩序列進行差異分析,個體間堿基不配對分布曲線呈明顯的單峰形被認為種群歷史有擴張現象;其二是通過Tajima’sD和Fu’sFs中性檢驗,D和Fs呈負值和差異顯著的P值被認為種群在歷史上有擴張跡象。中性檢驗結果顯示,4個湖泊種群的D和Fs均為負值,但只有洪澤湖和駱馬湖種群的中性檢測統計檢驗有顯著性差異(表5)。另外,堿基不配對分布曲線呈單峰形,均說明太湖新銀魚在歷史上可能發生過種群擴張。
3.3 太湖新銀魚種群遺傳結構
種群遺傳結構的研究不僅可以用于評價物種群體間的變異水平和不同地理群體之間的關系,還可以確定群體中的進化顯著單元和管理單元,以及制定資源的保護和管理策略[27]。遺傳分化系數Fst是群體間遺傳分化的重要參數之一,其值大小能反映群體間分化程度,Fst值為0~0.05時無分化,0.05~0.15為中度分化,0.15~0.25為高度分化[28]。本研究分子方差分析結果顯示,群體間遺傳分化系數Fst=0.768 43(P=0.000 0),表明在整個遺傳變異中群體間占76.84%,其余的遺傳變異來自于群體內,群體間具有程度較高的遺傳分化。進一步比較兩兩群體間的遺傳分化系數,結果表明,太湖和洪澤湖群體間及高郵湖和駱馬湖群體間遺傳分化較小,遺傳分化不顯著;太湖和洪澤湖群體與高郵湖和駱馬湖群體間遺傳分化較大,遺傳分化顯著。單倍型最小網絡進化圖也顯示太湖和洪澤湖群體聚為一個進化單元,高郵湖和駱馬湖群體聚為一個進化單元。
一般來說,魚類的遺傳分化格局往往與其分布的水系格局和地理距離密切相關[29-30]。從水系分布來看,太湖屬于長江水系,高郵湖、洪澤湖和駱馬湖則屬于淮河水系;從地理位置來看,太湖、高郵湖、洪澤湖和駱馬湖從南向北依次分布。由此可以得出,4個湖泊群體的遺傳結構關系與水系分布及地理距離不相符合。同樣,基于線粒體Cytb基因序列的太湖新銀魚和喬什新銀魚遺傳分布格局也得到類似結果,并且發現銀魚的遺傳格局與進化歷史有密切關系[9,12]。同樣,本研究中4個太湖新銀魚群體遺傳格局可能更多受進化歷史的影響。研究表明,銀魚起源于第三紀中期,進化過程中經歷了第四紀冰期和間冰期氣候的交替變化,導致銀魚棲息地數度隔離和連接,從而對銀魚的進化分化過程造成了重大影響[31]。基于Cytb基因的長江流域和淮河流域太湖新銀魚種群擴張分別發生在2.21萬年前和1.90萬年前[12],均早于發生在約1.8萬年前第四紀冰期最后一次大冰期[32]。因此可以推測,最后一次大冰期對太湖新銀魚種群產生很大影響,冰期期間太湖新銀魚生活在某個避難所內,隨著最后一次冰期結束,氣候變暖,海平面上升,太湖新銀魚種群向外擴散。在隨后的進化過程中逐漸產生不同的周單倍型,形成現今的遺傳結構模式。
3.4 太湖新銀魚種質資源保護
太湖新銀魚是我國特有的銀魚種類,具有較高的經濟價值和營養價值,曾是我國重要的漁業捕撈對象和出口創匯水產品。太湖新銀魚屬于典型的r-策略生物,生命周期只有1年,對環境變化比較敏感,種群易于波動。近年來,受過度捕撈、環境污染、棲息地破壞等多種不利因素的影響,太湖新銀魚資源已趨于枯竭,可持續發展受到嚴重威脅,因此,必須加強對太湖新銀魚資源的保護和管理。本研究基于COⅠ基因序列,調查了江蘇省4個湖泊太湖新銀魚群體的遺傳多樣性水平,分析了它們的遺傳結構關系,為制定相關的保護和管理措施提供了依據。根據研究結果,建議將太湖和洪澤湖種群及高郵湖和駱馬湖種群分別作為一個整體進行管理和保護;將提高太湖新銀魚遺傳多樣性作為重點。為了恢復太湖新銀魚資源,可以采取以下措施:1)控制水環境污染,保護棲息環境,為太湖新銀魚生活和繁殖提供良好的生態環境;2)嚴格控制捕撈強度,嚴禁使用帶有毀滅性破壞的電魚、炸魚等捕撈方法,禁止產卵期捕魚;3)加強太湖新銀魚人工繁殖技術研究,大力開展人工增殖放流,減少捕撈量;4)加強漁業資源和生態環境監測,掌握太湖新銀魚種群動態變化。
致謝:江蘇省太湖漁業管理委員會辦公室、江蘇省高寶-邵伯湖漁業管理委員會辦公室、江蘇省洪澤湖漁業管理委員會辦公室及江蘇省駱馬湖漁業管理委員會辦公室在采樣過程中給予很大的幫助,一并表示感謝!
