海州灣海洋牧場拖網漁獲物營養級分析
2020-07-01 05:07:08高世科于雯雯吳立珍路吉坤伏光輝
廣東海洋大學學報 2020年4期
張 碩,高世科,張 虎,于雯雯,吳立珍,路吉坤,伏光輝
海州灣海洋牧場拖網漁獲物營養級分析
張 碩1,高世科1,張 虎2,于雯雯2,吳立珍3,路吉坤4,伏光輝4
(1. 上海海洋大學海洋科學學院,上海 201306;2. 江蘇省海洋水產研究所,江蘇 南通 226007;3. 連云港市海域使用保護動態管理中心//4. 連云港漁業發展促進中心,江蘇 連云港 222002)
【目的】分析與對比2014/2018年海州灣同種生物的營養級特征。【方法】根據2018年9月海州灣海洋牧場區漁業資源調查,應用穩定同位素方法計算主要生物的δ13C、δ15N值及營養級,運用系統聚類分析劃分主要營養類群(Trophic Group, TG)。【結果】魚類、蝦蟹類、頭足類和貝類,營養級范圍分別為2.6 ~ 4.1、2.7 ~ 3.6、2.9 ~ 3.3和2.0 ~ 3.0。該海域的拖網漁獲物可分為3大類群:TG1包括大多數生物類群,由魚、蝦、蟹類和頭足類組成;TG2主要為高營養級消費者,包括斑尾刺鰕虎魚和四指馬鲅;TG3為初級消費者。【結論】2018年海州灣海洋牧場區域同種生物的營養級較2014年明顯變高,但也有個別種類的營養級出現不同程度的降低。
海州灣;海洋牧場;穩定同位素;營養級
水生生態系統中能量的流動和轉化對水環境的物質循環有著極其重要的作用[1]。為闡釋海洋生物間捕食、競爭和共生等關系,構建完整海域食物網、分析食物網結構和功能具有長足意義[1-3]。傳統的胃含物分析法雖能研究生物在一定時期內的食性變化,但耗時費力,隨著穩定同位素技術日臻成熟,利用碳(δ13C)、氮(δ15N)穩定同位素技術來分析海洋生物的營養功能和結構已成為海洋食物網研究工作中一種重要手段[5-6]。
在海洋食物網中,碳穩定同位素(δ13C)主要用于示蹤生物的食物來源,還能反映生物所在的海域差異,氮穩定同位素(δ15N)在生物體內逐層富集,可用于計算營養級[7]。基于碳氮穩定同位素技術,我國黃東海[8]、渤海[9]、膠州灣[10]、長江口[11]等沿海海域逐漸建立起食物網基礎構架。
海州灣屬于半開闊海域,生物資源量豐富,是我國重點保護的港灣之一[12]。近年來,由于航道建設、過度捕撈等一系列人為活動開展,海州灣海域環境受到不利影響。自2007年起,當地政府開始建設以人工魚礁為主的海洋牧場,用以生態環境修復和資源利用[13]。2014年張碩等[14]和2017年謝斌等[15]基于穩定同位素技術定量研究了海州灣海洋牧場營養基礎和群落營養結構的季節性變化。
為掌握不同年度海洋牧場生物營養結構變化情況,本研究通過碳、氮穩定同位素方法研究2018年海州灣拖網漁獲物營養級的狀況,對比分析海州灣海洋牧場2018年與2014年漁獲物營養級的差異,以及海洋牧場生態工程建設隨時間遞增對海域食物網結構的影響,以期為海州灣近海生態環境修復進一步開展提供參考。
1 材料與方法
1.1 樣品采集
所有生物樣品于2018年9月在海州灣海洋牧場區域內采集得到,站位設置范圍在34°52.849′N-34°56.117′ N, 119°13.641′ E-119°33.778′ E(圖1)。本次調查使用單船底拖網(規格125.32 m×59.1 m×36.0 m),平均拖時為30 min,船速為2.3 kn,共采集28種生物用于穩定同位素分析,其中魚類13種,頭足類3種,蝦蟹類8種,貝類共4種。具體數量見表1。
圖1 海州灣海洋牧場區域調查站位
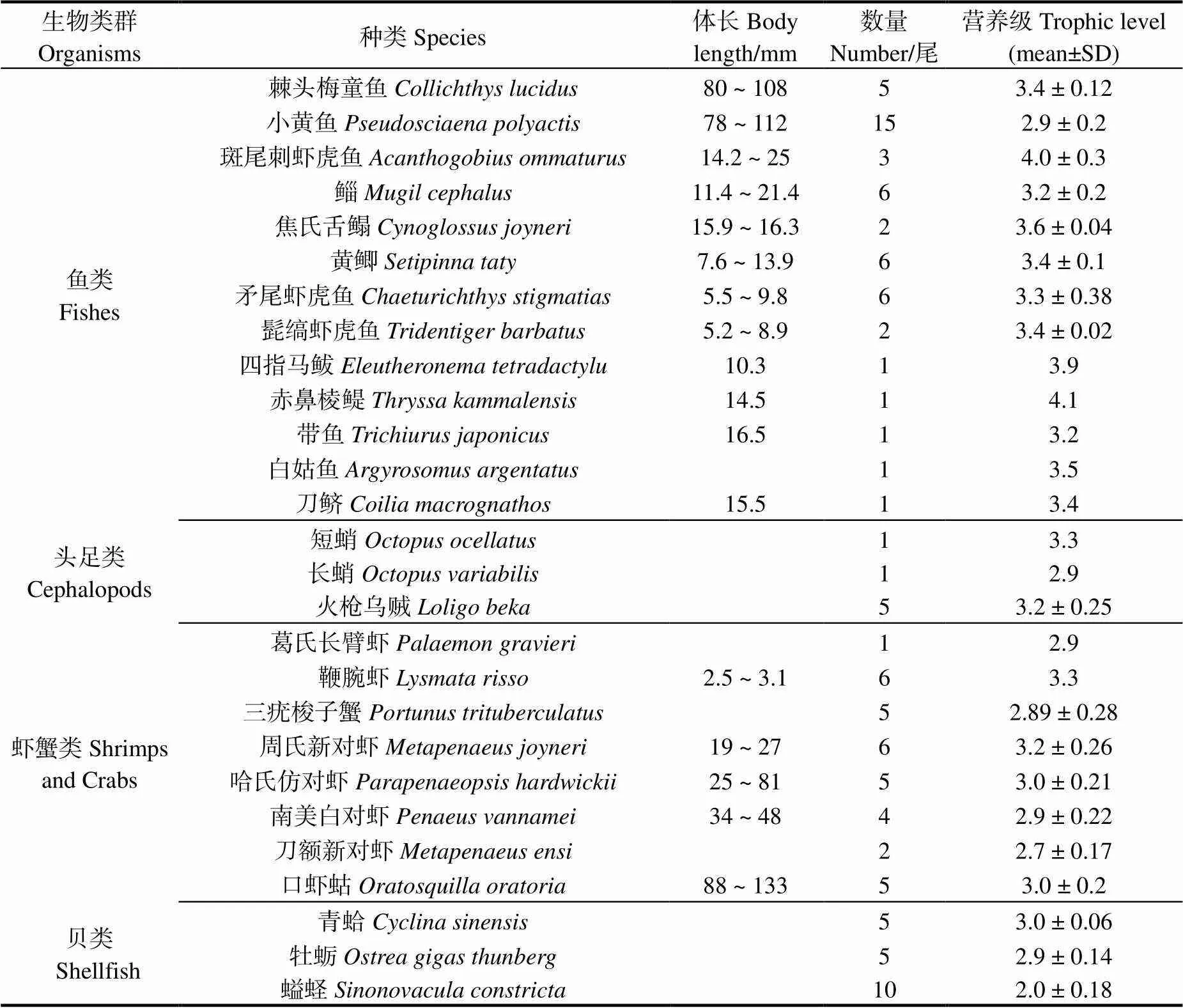
表1 海州灣海洋牧場區域拖網漁獲物的營養級
1.2 樣品處理及分析
所有生物樣品冷凍保存并運輸至上海海洋大學攝食生態實驗室進行室內分析。根據《海洋調查規范》[16]進行生物學基礎指標測定(表1),隨后取魚類背部白肌、蝦類腹部肌肉、蟹類第一螯足、雙殼類閉殼肌、腹足類斧足和頭足類胴部。所有樣品均在-44.6℃冷凍干燥后研磨備用。由于脂質含量只對肌肉組織的13C含量有影響,因此本研究未對肌肉組織進行去脂化處理[16]。
穩定同位素的分析在上海海洋大學漁業資源可持續開發平臺教育部重點實驗室分析測試平臺完成,使用德國vario元素分析儀和Isoprime100質譜儀測定碳、氮穩定同位素比值,分別以國際通用標準物質PeeDee石灰巖中的碳和大氣氮(N2)作為參考標準,結果以13C和15N(平均數±標準差)形式來表示,通過以下公式計算:
X=(sample/standard-1)×103, (1)
式中,X為13C或者15N;為13C/12C或者15N/14N的比值。
營養級的確定公式如下:
TL=[(15Nsample-15Nbaseline)/△15N]+2, (2)
式中,TL表示所計算生物的營養位置;15Nsample為系統消費者氮穩定同位素比值;15Nbaseline為該系統基線生物的氮穩定同位素比值,本研究選取縊蟶[14];△15N為1個營養級的富集度(Trophic Enrichment Factor),本研究沿用Post等△15N的均值3.4‰[18]。常數項2為1個基線生物的營養級。
1.3 數據處理
通過ArcGis 10.3進行站位圖的繪制;基于各類生物的13C和15N值的平均標準化歐氏距離(Standardised Euclidean distance),通過SPSS 25.0軟件進行組間連接系統聚類分析(Hierarchical Cluster Analysis);其余數據的處理、統計通過Excel 2019協助完成。
2 結果
2.1 漁獲物碳氮穩定同位素比值
2018年調查,海州灣海洋牧場區域拖網漁獲物種類的13值范圍在-21.71‰ ~ -16.20‰,平均值為(18.00 ± 1.19)‰;15N值范圍在(7.18‰ ~ 14.44)‰,平均值為(11.09 ± 1.62)‰(圖2)。
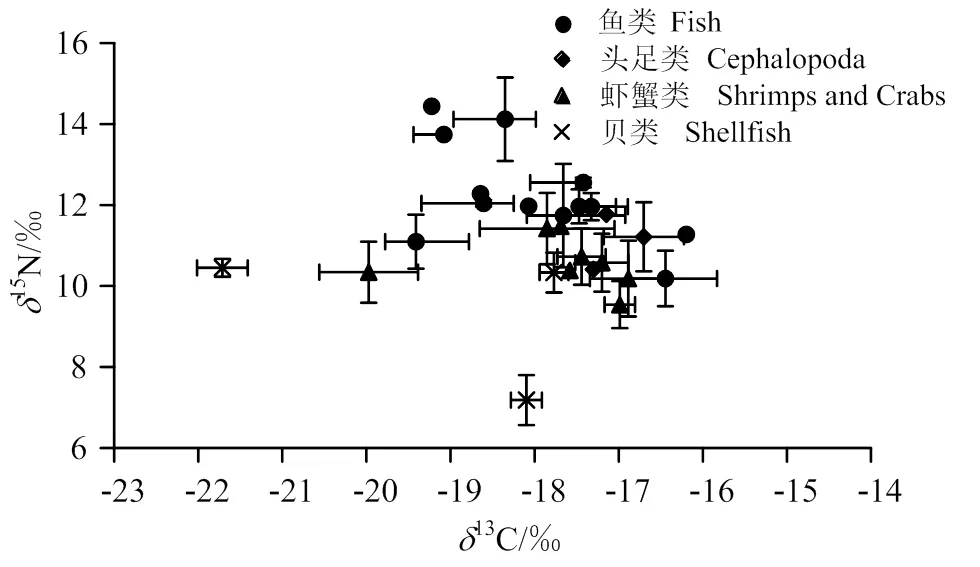
圖2 海州灣海洋牧場區域拖網漁獲物(Mean± SD)二維分布
魚類13C和15N值范圍分別為-19.41‰ ~ -16.20‰和10.18‰ ~ 14.44‰,跨度較大,分別為3.21‰和4.26‰;13C和15N值的平均值分別為(-17.99 ± 0.98)‰和(12.26 ± 1.17)‰。
頭足類13C和15N值范圍為-17.3‰ ~ -16.70‰和10.41‰ ~ 11.77‰,13C值跨度為0.60‰,15N值跨度為1.36‰;13C、15N平均值分別為(-17.05 ± 0.25)‰和(11.13 ± 0.56)‰。
蝦蟹類13C范圍為-19.97‰ ~ -16.89‰,平均值為(-17.70 ± 0.91)‰;15N值范圍為9.54‰~11.48‰,平均值(10.58 ± 0.60)‰。13C、15N值的跨度分別為3.08‰和1.94‰。
貝類的13C值范圍較廣,為-21.71‰ ~ -17.77‰,平均值為(-18.85 ± 1.66)‰;15N值較低,為7.18‰ ~ 10.45‰,平均值為(8.85 ± 1.54)‰。13C、15N值的跨度都較大,分別為3.94‰和3.27‰。
2.2 基于聚類分析劃分的不同營養類群
通過基于13C和15N值的標準化歐氏距離(Standardised Euclidean distance)的組間連接系統聚類分析可知,本次海州灣海洋牧場區域生物群落基本上可分為3大營養類群(Trophic Group)(圖3),TG1為第一營養類群,主要代表為棘頭梅童魚、小黃魚等魚類、周氏新對蝦、三疣梭子蟹等蝦蟹類和火槍烏賊、長蛸等頭足類;TG2主要為高級消費者,包括斑尾刺鰕虎魚、四指馬鲅等;TG3為營養級最低的初級消費者。

圖3 海州灣海洋牧場區域拖網漁獲物碳氮穩定同位素比值的聚類分析
2.3 營養級的確定與年間對比
根據營養級大小將海州灣海洋牧場區域拖網漁獲物主要劃分為4大類群(圖3),魚類的營養級范圍介于2.9(小黃魚)~ 4.1(赤鼻棱鳀)之間,其中營養級在3.0以下的只有小黃魚,有8種魚類集中在3.0 ~ 3.5之間,集中在3.5 ~ 4.1之間的魚類有4種,高于4.0的只有赤鼻棱鳀;蝦蟹類營養級在2.7(刀額新對蝦)~ 3.3(鞭腕蝦)范圍內變化,營養級在2.5 ~ 3.0之間的有5種,在3.0 ~ 3.5之間的有3種;頭足類營養級范圍為2.9(長蛸)~ 3.3(短蛸);貝類營養級范圍為2.0(螠蟶)~ 3.0(青蛤)。
如表2,2018年海州灣海洋牧場區域相同生物的營養級明顯高于2014年,其中,營養級差值最大的是口蝦蛄,為0.6個營養級,其次是棘頭梅童魚和火槍烏賊,小黃魚、鯔的營養級差為0.4,斑尾刺蝦虎魚和三疣梭子蟹的營養級差為0.3,剩余有53.3%的種類的營養級差集中在0.1 ~ 0.2之間,相差不大。

表2 2014年和2018年海州灣海洋牧場區域相同生物的數量和營養級比較
3 討論
3.1 海州灣拖網漁獲物的穩定同位素特征
本次海州灣海洋牧場區域調查拖網漁獲物的15N值出現由高到低的階梯式分布。魚類在生物樣品中所占比例較大,其中蝦虎魚科魚類屬于兇猛肉食性魚類,其15N值始終保持在較高水平[19]。棘頭梅童魚、小黃魚等石首魚科魚類的15N值與長江口海域同一體長規格的魚類相比,大致相符[11, 20],本研究赤鼻棱鳀、黃鯽的15N值變化較大,原因可能與其攝食較大比例底層蝦類有關[21]。焦氏舌鰨的15N值與其他海域同屬舌鰨相比偏小[8, 10],這與海域差異、調查時間及體長規格等因素有關。
系統聚類分析結果中,TG1中多數魚類、蝦蟹類屬于中級消費者范疇,有些蝦蟹類和頭足類的營養級甚至超過部分魚類,在其他海域也出現過這種情況[10],這是由于魚類捕食營養級較低的生物,導致營養級降低所致[22],這也是TG1中一些魚類與蝦類被歸為一類的原因。鯔在各大海域被視為中型捕食者,在本研究中與南美白對蝦同屬一個分類,可能原因是鯔的13C值較低,導致其與蝦類的食性相近。
3.2 影響海域食物網中營養級確定的主要因素
基線生物和△15N值是影響食物網中生物確定營養級的關鍵因素[23]。基線生物一般選取生長周期長、食性相對穩定的初級消費者、底棲動物或POM等,還需考慮基線生物的時空異質性,且對于生境結構復雜的海域,則可能需要多個基線生物以全面表征食物網結構[24]。縊蟶作為本研究的基線生物,其△15N值與2014年差異較小,且生活史和生長環境比較符合該海域基線生物選取的標準[14]。
△15N值用于定量分析生態系統中生物的營養級。根據15N值在生物體內富集規律的不同,所得到的△15N值也有所差異。一般而言,選用水生生態系統生物的△15N值作為研究對象,且排泄物是氨氮的生物比排泄物是尿素的生物的△15N值小,能夠減小誤差[25]。Hussey等[26]提出,△15N值并非一個定值,使用一定范圍內的△15N值(scaled △15N framework)能提高食物網構建的精準度,意味著以往食物網結構被簡化。本研究沿用Post等[14]的△15N值3.4‰,亦是為了與2014年的研究一致,減小計算營養級的誤差。
此外,生物營養級還與體長因素密不可分[27]。Ji等[20]研究表明,小黃魚的15N值隨著體長增加而增大,蔡德陵等[8]研究證明帶魚在不同生命階段的15N值與其體長呈現正相關性;謝斌等[14]也通過對比鱸魚體長與其15N值的關系應證了這一點。這是由于有些生物在生活史中存在食性轉變現象,從幼體攝食營養級較低的食物轉變為成體攝食營養級較高的食物,導致營養級產生差異[28]。
3.3 海州灣海洋牧場不同年份間的比較
報道[29]表明,海洋牧場中人工魚礁的建設會使海域內的食物網結構發生改變,且隨年份遷移,差異變化更加明顯。本研究和2014年所設置的站位均在海州灣海洋牧場區域內,且隨著時間推移,同種生物的營養級較2014年有明顯上升趨勢,但也有個別種類營養級發生不同程度降低的現象。
林會潔等[29]研究表明,口蝦姑在人工魚礁區域比在貝類養殖區、海藻養殖區低了0.1 ~ 0.3個營養級,盡管海洋牧場的生態工程建設對海洋生物營養結構起到一定改善作用,豐富了海洋生物的種類和數量,但區域內存在激烈的生物間競爭和明顯生態位重疊,即“優勝劣汰”原則,因而導致有些底棲生物種類由于數量增多會因短期內食物來源匱乏而出現營養級降低的現象。這一結論應證了本研究口蝦姑營養級降低的可能原因。同時,也能夠解釋本研究蝦蟹類的營養級在2個年份間出現不同程度下降的現象。而火槍烏賊的食性以甲殼類為主,其中口蝦蛄、葛氏長臂蝦占比為4.7%和1.3%[30],由于生物餌料組成減少或者消失,致使其強制性改變食性或食源,以更低營養級的餌料為食,因此這可能是火槍烏賊營養級下降的主要原因。
小黃魚是主要以魚類、甲殼類為食的雜食性魚類,口蝦姑、葛氏長臂蝦是其食物之一[28]。本研究小黃魚與2014年相比下降了0.4個營養級,在體長規格相差不大的情況下,除考慮可能的食性轉變對其營養級的影響,還需進一步分析其潛在碳源影響。謝斌等[15]研究表明,小黃魚的潛在碳源以浮游植物為主,而近年來,海洋牧場的建立減少了水體富營養化和赤潮發生率,改善了水環境質量,使得海州灣夏季浮游植物濃度逐年波動幅度較大[31],導致小黃魚的潛在碳源出現不可預測的變化,這可能是小黃魚營養級下降的另一個潛在因素。
4 結語
海洋牧場生態工程建設對食物網結構的影響是多方面的,一方面增加了物種多樣性,增大了食物網結構的復雜程度;另一方面,在改善生物生存環境的同時,出現了高營養級生物數量增多、低營養級生物數量減少的現象,以致于部分生物種類的營養級降低。本研究僅從2年份間生物的碳氮穩定同位素值來界定海州灣海洋牧場建設對生態系統的影響,顯然不夠全面,在將來研究中,可將水環境因素(pH、光照、溫度、鹽度等)結合生態系統評估系統引入,綜合考量海洋牧場建設帶來的生態效益,也能為海州灣近海環境保護和漁業資源恢復問題拓展相應的理論參考。
[1] ZADEREEV E S, GULATI R D, CAMACHO A. Biological and ecological features, trophic structure and energy flow in meromictic lakes[M]//Ecology of Meromictic Lakes. Cham: Springer International Publishing, 2017: 61-86.
[2] BUMPERS P M, ROSEMOND A D, MAERZ J C, et. al. Experimental nutrient enrichment of forest streams increases energy flow to predators along greener food-web pathways[J]. Freshwater Biology, 2017. 62(10): 1794-1805.
[3] B?NARU D, HARMELIN-VIVIEN M. Trophic links and riverine effects on food webs of pelagic fish of the north-western Black Sea[J]. Marine and Freshwater Research, 2009, 60(6): 529.
[4] JOHANNSSON O E, LEGGETT M F, RUDSTAM L G, et al. Diet ofin Lake Ontario as revealed by stable isotope and gut content analysis[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2001, 58(10): 1975-1986.
[5] CABANA G, RASMUSSEN J B. Comparison of aquatic food chains using nitrogen isotopes[J]. Proceedings of the National Academy of Sciences of the United States of America, 1996, 93(20): 10844-10847.
[6] PRECIADO I, CARTES J E, PUNZóN A, et al. Food web functioning of the benthopelagic community in a deep-sea seamount based on diet and stable isotope analyses[J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2017, 137: 56-68.
[7] WADA E, TERAZAKI M, KABAYA Y, et. al.15N and13C abundances in the Antarctic Ocean with emphasis on the biogeochemical structure of the food web., 1987, 34: 829-841.
[8] 蔡德陵, 李紅燕, 唐啟升, 等. 黃東海生態系統食物網連續營養譜的建立: 來自碳氮穩定同位素方法的結果[J]. 中國科學C輯: 生命科學, 2005, 35(2): 123-130.
[9] QU P, WANG Q X, PANG M, et al. Trophic structure of common marine species in the Bohai Strait, North China Sea, based on carbon and nitrogen stable isotope ratios[J]. Ecological Indicators, 2016, 66: 405-415.
[10] 麻秋云, 韓東燕, 劉賀, 等. 應用穩定同位素技術構建膠州灣食物網的連續營養譜[J]. 生態學報, 2015, 35(21): 7207-7218.
[11] 閆光松, 張濤, 趙峰, 等. 基于穩定同位素技術對長江口主要漁業生物營養級的研究[J]. 生態學雜志, 2016, 35(11): 3131-3136.
[12] 張秀英, 鐘太洋, 黃賢金, 等. 海州灣生態系統服務價值評估[J]. 生態學報, 2013, 33(2): 640-649.
[13] 吳立珍, 吳衛強, 陸偉, 等. 海州灣生態環境修復的探索實踐與展望: 江蘇省海洋牧場示范區建設[J]. 中國水產, 2012(6): 35-37.
[14] 張碩, 謝斌, 符小明, 等. 應用穩定同位素技術對海州灣拖網漁獲物營養級的研究[J]. 海洋環境科學, 2016, 35(4): 507-511.
[15] 謝斌, 李云凱, 張虎, 等. 基于穩定同位素技術的海州灣海洋牧場食物網基礎及營養結構的季節性變化[J]. 應用生態學報, 2017, 28(7): 2292-2298.
[16] 中華人民共和國國家質量監督檢驗檢疫總局, 中國國家標準化管理委員會. GB/ T 12763. 6-2007 海洋調查規范第6 部分: 海洋生物調查. 北京: 中國標準出版社, 2007
[17] SKINNER M M, MARTIN A A, MOORE B C. Is lipid correction necessary in the stable isotope analysis of fish tissues[J]. Rapid Communications in Mass Spectrometry, 2016, 30(7): 881-889.
[18] POST D M. Using stable isotopes to estimate trophic position: models, methods, and assumptions[J]. Ecology, 2002, 83(3): 703-718.
[19] NEVES L M, TEIXEIRA T P, ARAúJO F G. Structure and dynamics of distinct fish assemblages in three reaches (upper, middle and lower) of an open tropical estuary in Brazil[J]. Marine Ecology, 2011, 32(1): 115-131.
[20] JI W, CHEN X, JIANG Y, et. al. Trophic ecology of small yellow croaker (Larimichthys polyactis Bleeker): stable carbon and nitrogen isotope evidence[J]. Chinese Journal of Oceanology and Limnology, 2011, 29(5): 1033-1040.
[21] 王田田, 呂振波, 李凡, 等. 基于穩定碳氮同位素的萊州灣4種鳀鯡科魚類營養級研究[J]. 中國水產科學, 2013, 20(5): 1076-1085.
[22] MALEK A J, COLLIE J S, TAYLOR D L. Trophic structure of a coastal fish community determined with diet and stable isotope analyses[J]. Journal of Fish Biology, 2016, 89(3): 1513-1536.
[23] JUN X U, ZHANG M A, XIE P. Variability of stable nitrogen isotopic baselines and its consequence for trophic modeling[J]. Journal of Lake Sciences, 2010, 22(1): 8-20.
[24] 貢藝, 陳玲, 李云凱. 海洋生態系統穩定同位素基線的選取[J]. 應用生態學報, 2017, 28(7): 2399-2404.
[25] CAUT S, ANGULO E, COURCHAMP F. Variation in discrimination factors (Δ15N and Δ13C): the effect of diet isotopic values and applications for diet reconstruction[J]. Journal of Applied Ecology, 2009, 46(2): 443-453.
[26] HUSSEY N E, MACNEIL M, MCMEANS B C, et al. Rescaling the trophic structure of marine food webs[J]. Ecology Letters, 2014, 17(2): 239-250.
[27] VILLAMARíN F, JARDINE T D, BUNN S E, et al. Body size is more important than diet in determining stable-isotope estimates of trophic position in crocodilians[J]. Scientific Reports, 2018, 8: 2020.
[28] XUE Y, JIN X, ZHANG B, et. al. Seasonal, diel and ontogenetic variation in feeding patterns of small yellow croaker in the central Yellow Sea[J]. Journal of Fish Biology, 2005, 67(1): 33-50.
[29] 林會潔, 秦傳新, 黎小國, 等. 柘林灣海洋牧場不同功能區食物網結構[J]. 水產學報, 2018, 42(7): 1026-1039.
[30] 楊紀明. 渤海無脊椎動物的食性和營養級研究[J]. 現代漁業信息, 2001, 16(9): 8-16.
[31] 李大鵬, 張碩, 石一茜, 等. 海州灣海洋牧場浮游植物群落年際變A F G. Structure and dynamics of distinct fish assemblages in three reaches (upper, middle and lower) of an open tropical estuary in Brazil[J]. Marine Ecology, 2011, 32(1): 115-131.
Time-scale Variation and Trophic Level Analysis of Stable Isotope of Major Organisms in Haizhou Bay
Zhangshuo1, Gaoshi-ke1, Zhanghu2, Yuwen-wen2,, Wuli-zhen3, Lu ji-kun4, Fu guang-hui4
(1.,,201306,; 2.,226007,,3.//4.,222002,)
【Objective】The trophic level of the same organisms in the same area, 2014 were compared and analyzed. 【Method】According to the fishery resource investigated in Haizhou Bay, 2018, theδ13C, δ15Nvalue and trophic level of major organisms were calculated using stable isotope method. The trophic groups (TGs) were classified by hierarchical cluster analysis. 【Result】Fish, crustacean, cephalopods and shellfish was divided in food web, in which the trophic level ranged from 2.6 to 4.1, 2.7 to 3.6, 2.9 to 3.3 and 2.0 to 3.0, respectively. TG1 (Trophic Group 1) included small-sized fishes, shrimps, crabs and a few cephalopods; TG2 consists mainly of advanced consumers like,and primary consumers in TG3. 【Conclusion】The trophic level of same organsims in 2018 was remarkedly higher than that in 2014 in Haizhou Bay. However, the trophic level of several species reduced at different degrees.
Haizhou bay; Sea ranching; Stable isotope; Trophic level
S 931.1
A
1673-9159(2020)04-0132-07
10.3969/j.issn.1673-9159.2020.04.018
2019-12-10
江蘇省自然科學基金青年基金(BK20170438);南通市科技計劃項目(JC2018014)
張碩(1976-),男,博士,教授,主要研究方向為近海生態環境修復。E-mail: s-zhang@shou.edu.cn
于雯雯(1983-),男,博士,副研究員,主要研究方向為海洋生態方面。E-mail: jshyyww@163.com
張碩,高世科,張虎,等. 海州灣海洋牧場拖網漁獲物營養級分析[J].廣東海洋大學學報,2020,40(4):132-138.
(責任編輯:劉嶺)
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
今日農業(2021年11期)2021-08-13 08:53:34
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
作文周刊·小學一年級版(2017年17期)2017-06-27 19:32:55
