黃瓜實生苗與嫁接苗生長發育及光合熒光特性對自毒脅迫的響應差異
2020-07-08 09:47:18馬寧陳碧楊華劉威黃興昌肖雪梅頡建明
中國瓜菜 2020年6期
馬寧 陳碧 楊華 劉威 黃興昌 肖雪梅 頡建明


摘 要: 采用水培方式,以黃瓜嫁接苗與實生苗為試材,外源肉桂酸(CA)模擬自毒脅迫,分別從幼苗的生長發育、根系形態和光合熒光特性等方面,對比研究以黑籽南瓜為砧木的嫁接苗與黃瓜實生苗對自毒脅迫響應差異的生理機制。結果表明,第6天時,CA脅迫下黃瓜實生苗(NG-T)的株高和葉面積比對照(NG-CK)分別降低了32.96%和33.41%,而嫁接苗的株高、莖粗、葉面積受抑制程度輕,且長勢明顯優于實生苗;0.5 mmol·L-1 CA對黃瓜嫁接苗的根長和根尖數的抑制率為6.95%和7.30%,僅為黃瓜實生苗的18.66%和16.37%;CA脅迫下,嫁接能夠穩定維持黃瓜幼苗體內的葉綠素含量;嫁接苗的蒸騰速率(Tr)、凈光合速率(Pn)、氣孔導度(Gs)、Fv/Fm、Fv/Fo和非光化學淬滅(NPQ)均高于實生苗,分別高出10.87%、156.86%、78.33%、5.58%、17.59%和38.07%。說明嫁接可促進黃瓜植株的生長,減輕自毒物質對光合系統的損傷,從而提高對自毒脅迫的耐受性。
關鍵詞: 黃瓜;肉桂酸;嫁接;植株形態;光合熒光特性
中圖分類號:S642.2? ? 文獻標志碼:A? ? 文章編號:1673-2871(2020)06-017-07
Abstract: In the experiment, hydroponics was used to study the difference physiological mechanism of the non-grafted and rootstock grafted cucumber seedling in response to autotoxic stress, using exogenous cinnamic acid(CA)to simulate autotoxic stress. The growth and development of seedlings, root morphology and photosynthetic fluorescence characteristics were compared between non-grafted and rootstock grafted cucumber. The results showed that on day 6(d), plant height and leaf area of non-grafted cucumber seedlings(NG-T)under cinnamic acid stress were 32.96% and 33.41% lower than those of control(NG-CK), while plant height, stem diameter and leaf areas of rootstock grafted seedlings were slightly inhibited, and growth vigor was significantly better than that of non-grafted seedlings. The inhibition rates of root length and root tip number of grafted seedlings under 0.5 mmol L-1 CA treatment were 6.95% and 7.30%, respectively, only 18.66% and 16.37% of the non-grafted cucumber seedlings, which indicated the resistance of grafted seedlings to CA stress was stronger. Under CA stress, grafting could keep the chlorophyll content of cucumber seedlings stable. The transpiration rate(Tr), net photosynthetic rate(Pn), stomatal conductance(Gs), Fv/Fm, Fv/Fo and non-photochemical quenching(NPQ)of grafted seedlings were increased than those of non-grafted seedlings by 10.87%, 156.86%, 78.33%, 5.58%, 17.59% and 38.07%, respectively. The results showed that grafting could promote the growth of cucumber plants, inhibit the damage of photosynthetic system caused by autotoxic substances, and improve the tolerance to autotoxic stress.
Key words: Cucumber; Cinnamic acid; Grafting; Plant morphology; Photosynthetic fluorescence characteristics
黃瓜(Cucumis sativus L.)是葫蘆科一年生蔓性植物,生長周期短,市場需求量大,其設施栽培面積也在日益增大。但栽培中連作障礙普遍發生,致使黃瓜產量銳減和品質下降,同時抗病蟲害的能力減弱,嚴重制約了黃瓜設施栽培的可持續發展。前人研究表明,導致連作障礙的因素主要有3個方面,一是根系分泌自毒物質,二是土壤營養失衡,三是土傳致病菌的積累,其中植物自身釋放的有毒物質可影響根系對礦質元素的吸收,增加致病菌的種類和數量[1-2],是導致連作障礙的重要因素。
許多蔬菜作物根系可分泌出酚酸類自毒物質,影響正常的生理代謝。黃瓜的自毒作用是由苯基羧酸類化合物引起的,包括苯甲酸和肉桂酸及其衍生物[3],這些自毒物質通過影響黃瓜幼苗的離子吸收和光合作用,抑制黃瓜生長,且具有劑量效應[4];同時會造成黃瓜根系氧化脅迫,引起膜質過氧化和質膜H+-ATPase活性的下降[5];促進枯萎病的發生。Ding等[6]報道指出,黃瓜根系分泌的苯丙烯酸對自身存在毒害作用,當質量分數超過0.05 mg·g-1時,可顯著抑制黃瓜幼苗生長、根系活力與微生物活動,且濃度越高,抑制作用越強。同樣地,大量外源添加自毒物質試驗證實,自毒物質通過抑制作物根系生長、葉綠素合成及離子吸收能力,從而影響作物的生長發育,在黃瓜、豌豆、茄子、番茄等[7-10]作物中均有報道。
瓜類嫁接栽培在農業生產中已表現出抗病、抗逆、高產、優質的綜合優良性狀,得到了生產者的普遍認可。有研究證實,作為黃瓜嫁接砧木的黑籽南瓜不但不易產生自毒作用,其生長有時反而被其它瓜類的根系分泌物所促進[11]。陳紹莉等[12]通過添加外源自毒物質肉桂酸和香草醛,探討了茄子實生苗和嫁接苗對自毒脅迫的生理響應差異,發現嫁接茄子的根系活力、抗氧化酶活性和滲透調節能力均高于非嫁接茄子。筆者基于嫁接技術在減輕設施黃瓜連作障礙中的應用,采用外源肉桂酸模擬自毒脅迫,以黃瓜嫁接苗和實生苗為試材,分別從生長發育、根系形態和光合作用等方面闡述嫁接提高黃瓜抗自毒脅迫的生理機制,為生產中應用嫁接技術克服黃瓜連作障礙提供科學依據。
1 材料與方法
1.1 材料
試驗于2017年10月至2018年4月在甘肅農業大學園藝學院蔬菜栽培生理實驗室進行。接穗:‘津研四號黃瓜,天津市宏豐蔬菜研究有限公司。砧木:黑籽南瓜,云南農作資源開發研究所。
1.2 試驗設計與方法
選取健壯飽滿且形態一致的黃瓜種子溫湯浸種后,置于28 ℃人工氣候箱黑暗條件下催芽。當黃瓜子葉大部分露出或完全褪掉種皮時,移入預先準備好的1 L圓臺形水培盒(底徑12.5 cm,頂徑15.5 cm,高10 cm)內,采用日本山崎黃瓜專用營養液水培,在人工氣候箱中進行。培養條件:溫度25 ℃/19 ℃(晝/夜),白天光照強度為256 μmol·m-2·s-1,光周期14 h/10 h(晝/夜),濕度75%,營養液的電導率和pH分別為1.20 mS·cm-1、6.5。試驗期間每2 d更換1次營養液。當砧木(黑籽南瓜)第1片真葉露心,接穗(黃瓜)子葉完全展開時采用插接法進行嫁接。待幼苗長至2葉1心時,選取長勢一致的黃瓜嫁接苗(RG)與實生苗(NG),營養液中添加0.5 mmol·L-1 CA,以不加CA為對照。共如下4個處理:RG-CK(嫁接苗+營養液)、RG-T(嫁接苗+含0.5 mmol·L-1CA的營養液)、NG-CK(實生苗+營養液)、NR-T(實生苗+含0.5 mmol·L-1CA的營養液)。試驗采用隨機區組設計,每處理3個重復,每重復9株(每個水培盒種植3株)。CA采用無水乙醇助溶,濃度控制在0.1%(V/V),對照營養液中加入同等量的無水乙醇。分別于處理的0、2、4和6 d,測定不同處理黃瓜實生苗和嫁接苗的生長發育和光合特性,于處理的第6天,取樣測定幼苗生物量、根系形態和葉綠素熒光等參數。
1.3 測定指標與方法
1.3.1 植株形態的測定 用直尺測量根莖連接處到生長點的高度作為株高;用游標卡尺測定子葉下1 cm處的莖粗;測量相同葉位功能葉的長與寬,根據公式計算總葉面積,葉面積=0.743×長×寬[13]。
1.3.2 根系形態的測定 用蒸餾水將待測根洗凈,放入透明塑料盤中,加蒸餾水至根系完全浸沒,并將其充分鋪展,使用根系掃描儀(EPSON Scan,Canada)進行掃描,WinRHIZO Pro LA2400軟件分析根系總長度、表面積、根體積和根尖數。
1.3.3 植株生物量的測定 用蒸餾水把黃瓜幼苗洗凈,隨后用濾紙將植株表面的水分擦干,分為地上部與地下部,分別測定鮮質量,然后置于烘箱105 ℃殺青30 min,再調至80 ℃烘干至恒重。
1.3.4 葉綠素含量的測定 于處理6 d后,各處理隨機采取黃瓜幼苗從上向下數第2片功能葉。將葉片剪碎后混勻,取0.1 g放入25 mL具塞試管中,注入10 mL 80%的丙酮溶液,黑暗下浸泡提取,每4 h震蕩1次,待葉片呈白色時,在440 nm、663 nm和645 nm處用UV-1800型分光光度計進行比色。使用公式Ca=12.71×A663-2.59×A645;Cb=22.88×A645-4.67×A663;Ca+b=20.29×OD645+8.02×OD663算得葉綠素a、葉綠素b含量和葉綠素a+b含量。
1.3.5 光合參數的測定 分別在處理的第0、2、4和6 d采用CIRAS-2型便攜式光合儀(PP-system, UK),測定黃瓜幼苗從上向下數第2片功能葉的凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)及蒸騰速率(Tr)。設置測定條件,內置LED光源的光量子通量密度(PFD):1 000 μmol·m-2·s-1,環境CO2濃度:380 μmol·mol-1,葉片溫度:25 ℃,相對濕度:75%。
1.3.6 葉綠素熒光參數的測定 于處理第6天使用調制葉綠素熒光成像系統(MAXI Imaging-PAM,Walz,Effeltrich,Germany)測定黃瓜幼苗從上向下數第2片功能葉的葉綠素熒光參數。測定前,先將黃瓜幼苗進行30 min暗適應,使其反應中心處于完全開放狀態,剪下完全展開的第2片功能葉,平鋪在調制葉綠素熒光成像系統的測定平臺上,在25 ℃下開始測定葉片的Fv/Fm、Fv/Fo、ΦPSII、NPQ和qP等葉綠素熒光參數。
1.3.7 快速光響應曲線的測定 在MAXI Imaging-PAM的Light Curve窗口設置光合有效輻射強度梯度依次為0、21、56、111、186、281、396、531、701、926、1 251 μmol·m-2·s-1,相鄰光照強度之間間隔20 s,由此可以得到PSII相對電子傳遞速率(rETR)隨光強PAR增加的變化趨勢。
1.4 數據處理
使用Excel 2010軟件進行數據整理及制圖。采用IBM SPSS Statistics 22.0統計分析軟件對數據進行獨立樣本t檢驗,顯著性檢驗水平為P<0.05。
2 結果與分析
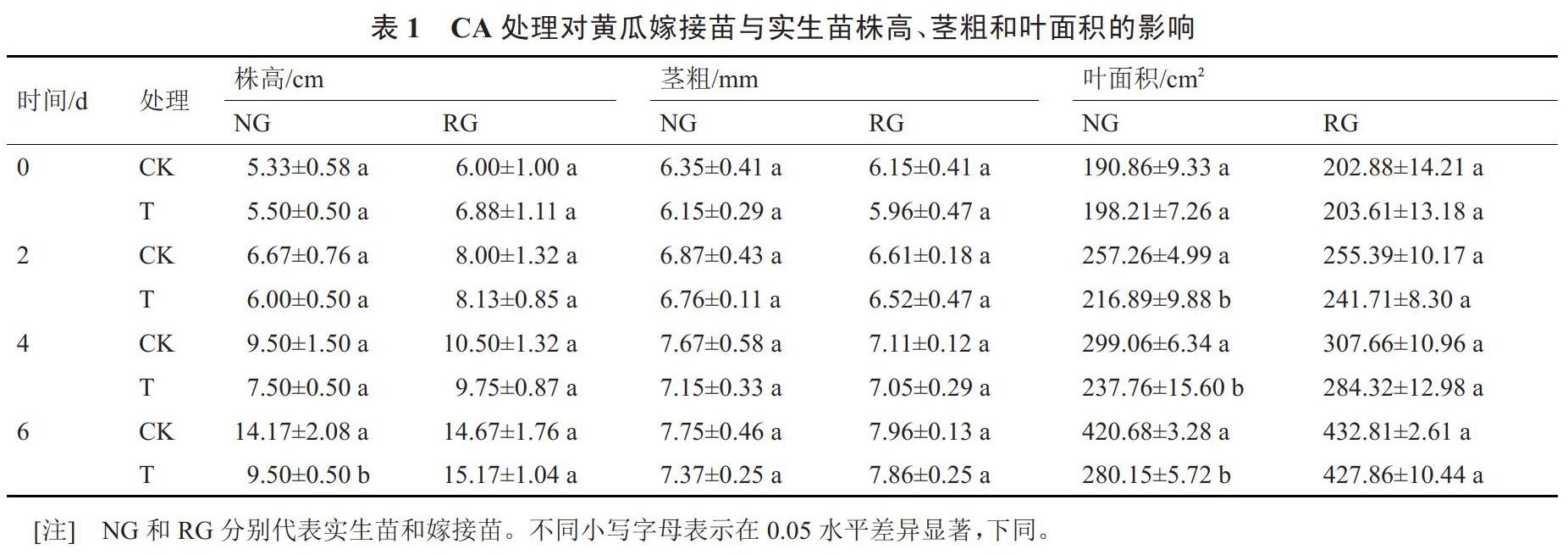
2.1 CA處理對黃瓜嫁接苗與實生苗生長發育的影響
由表1可以看出,0.5 mmol·L-1CA處理對NG幼苗生長有很強的抑制作用,在第6天時,NG-T的株高表現為被顯著抑制的現象,NG-T的株高較NG-CK降低32.96%。但NG莖粗并沒有被CA脅迫顯著抑制。同上,第2天時,NG-T幼苗的葉面積比NG-CK顯著減小15.69%。隨著處理時間的延長,抑制作用越明顯,在第6天時,NG-T的葉面積比NG-CK減小33.41%。而對于嫁接苗來說,0.5 mmol·L-1CA處理對幼苗株高、莖粗和葉面積均無顯著影響,且嫁接苗的各項形態指標均優于實生苗。說明嫁接對CA脅迫有很好的緩解作用。
2.2 CA處理對黃瓜嫁接苗與實生苗根系形態的影響
由表2可知,0.5 mmol·L-1CA處理顯著減少了黃瓜實生苗的根長、根尖數和根表面積,分別比對照降低37.24%、45.06%和6.12%;而根體積卻顯著增加40.19%。說明CA脅迫會對黃瓜根系產生很強的抑制作用。反觀RG-CK與RG-T,0.5 mmol·L-1CA處理會使嫁接苗的各項根系指標下降,但差異不顯著。說明嫁接苗對CA脅迫有很好的耐受性。嫁接砧木黑籽南瓜的黃瓜比實生黃瓜根系更發達,其根長、根表面積和根尖數分別是實生黃瓜的2.08倍、1.53倍和4.47倍,因此具有更強大的養分吸收能力以及對逆境的抵抗能力。
2.3 CA處理對黃瓜嫁接苗與實生苗生物量的影響
如圖1-A所示,0.5 mmol·L-1CA處理顯著降低了黃瓜實生苗的鮮物質積累量,NG-T的地上部和地下部的鮮物質積累量比NG-CK分別降低22.67%和28.93%,圖1-C顯示,0.5 mmol·L-1CA處理對其干物質量積累的影響沒有達到顯著水平。同時,由圖1-B、1-D可知,CA脅迫對RG地上和地下部的干、鮮物質積累量的影響都沒有表現出顯著差異。說明CA處理可能會引起水分脅迫進而抑制植株生長,而嫁接可以緩解甚至解除CA處理引起的水分脅迫,從而保證植株的正常生長發育。
2.4 CA處理對黃瓜嫁接苗與實生苗葉綠素含量的影響
由圖2-A可知,0.5 mmol·L-1CA處理可顯著降低黃瓜實生苗葉片葉綠素含量,NG-T的葉綠素a、葉綠素b和葉綠素a+b較NG-CK分別降低35.86%、47.68%和39.78%。圖2-B中,CA脅迫對RG葉綠素含量的影響無顯著差異,表明CA脅迫通過影響實生苗葉片光合色素的積累,進而影響植株的光合強弱,而嫁接可有效緩解CA脅迫對植株光合色素合成的抑制。
2.5 CA處理對黃瓜嫁接苗與實生苗光合特性的影響
由表3可以看出,0.5 mmol·L-1CA處理可顯著降低黃瓜實生苗的Tr、Gs和Pn,較對照分別降低了50.65%、49.55%和84.35%。在CA脅迫下,RG-T的Tr和Gs均降低,較RG-CK分別降低了7.96%和40.80%,而Pn卻沒有降低。與Tr、Gs和Pn不同的是,CA處理后,NG-T的Ci與對照相比顯著升高了34.77%;而RG-T與RG-CK的Ci無顯著差異,說明嫁接可有效緩解CA對黃瓜幼苗光合作用的脅迫。
2.6 CA處理對黃瓜嫁接苗與實生苗熒光參數的影響
光反應中心PSII的原初光能轉化效率以最大光化學效率(Fv/Fm)表示,PSII實際光化學效率(ΦPSII)則反映吸收的光量子供給PSII反應中心的效率及開放反應中心的比例。由表4可以看出,0.5 mmol·L-1CA處理可以降低NG的NPQ,顯著降低其Fv/Fm、Fv/Fo、ΦPSII和qP,分別比對照降低了4.27%、8.74%、21.07%和21.84%。而RG-T的NPQ和qP高于RG-CK,說明一定濃度CA脅迫時,嫁接會提高黃瓜葉片光合作用活性及光保護能力。對比NG-CK與RG-CK發現:RG-CK的Fv/Fm、Fv/Fo和NPQ均高于NG-CK,分別高出1.20%、8.32%、23.66%。RG-CK與RG-T之間各參數無顯著差異。
2.7 CA處理對黃瓜嫁接苗與實生苗快速光響應曲線的影響
由圖3可知,不同處理的黃瓜葉片隨光合有效輻射強度(PAR)的增加,其相對電子傳遞速率(rETR)變化趨勢保持一致,即先增加然后在光飽和狀態下達到穩定。從圖3-A可以看出,NG-CK的相對電子傳遞速率明顯高于NG-T,而RG-CK與RG-T差異不明顯(圖3-B)。在光飽和狀態下,RG-CK的電子傳遞速率較RG-T提高2.26%,較NG-CK提高8.32%,較NG-T提高22.09%,表明嫁接可以有效緩解CA處理對黃瓜幼苗葉片相對電子傳遞速率的抑制作用。
3 討論與結論
自毒作用是一個農業中普遍存在的現象,并且是造成連作障礙的主要因素之一[14],其作用機理復雜,植物在接觸、吸收化感物質后,導致生理生化過程紊亂。嫁接栽培在農業生產中已表現出抗病、抗逆、高產、優質的綜合優良性狀,在瓜類和茄果類園藝產品中得到廣泛應用[15]。有研究表明,嫁接不僅可以防治土傳病害,還可以提高產量、增強株苗的抗逆性,有效緩解黃瓜自毒造成的非生物脅迫[16]。本試驗結果也表明0.5 mmol·L-1 CA處理后的黃瓜嫁接苗的光合生理特性及生物量的積累均優于同處理下的黃瓜實生苗。
前人研究表明,黃瓜和黑籽南瓜對外源CA的響應存在顯著差異,外源CA處理對黃瓜植株生長產生抑制作用,而對黑籽南瓜植株生長無明顯影響[17]。本試驗對不同處理下黃瓜嫁接苗與實生苗的生長指標、根系形態及干物質的積累量進行測定,發現添加外源CA后,非嫁接黃瓜植株的株高、莖粗、葉面積增長及根系發育均低于對照植株,即外源CA對黃瓜幼苗的生長有很大的負面影響;而黃瓜嫁接苗的抑制程度較小。此結果與陳紹莉等[12]的一致,其研究表明較高濃度的CA(0.5~4.0 mmol·L-1)會顯著降低自根茄的株高、莖粗及鮮樣質量;較高濃度的CA(1~4 mmol·L-1)仍能促進嫁接茄的生長,只是促進效應有所減弱。
肖春蘭等[18]研究發現,黃瓜的Tr、Pn、Ci和Gs在其根系浸提液、分泌物及肉桂酸和苯甲酸類衍生物的影響下均有不同程度的降低。導致光合作用下降的因素有氣孔和非氣孔因素,如果氣孔限制因素占主導地位,在Pn和Gs下降的同時,Ci也相應下降;如果非氣孔限制因素占主導地位,則Pn和Gs下降的同時,Ci卻上升[19]。筆者發現,在不同濃度CA處理6 d后,黃瓜幼苗葉片Pn、Tr和Gs顯著降低,而Ci則上升,表明是非氣孔因素阻礙了CO2的利用,導致胞間CO2的積累,從而影響其光合能力,這與Allen和Ort研究一致[20]。賈聯紅[21]研究表明用嫁接西瓜莖葉、自根西瓜的根和莖葉的水浸液處理西瓜幼苗,當濃度為10 g·L-1時,Pn、Gs和Ci開始變小,隨濃度的增高,Pn、Gs和Ci降低,表明西瓜殘體水浸液通過影響Pn、Gs和Ci來阻礙西瓜幼苗的光合作用。本試驗中0.5 mmol·L-1 CA 處理后的黃瓜嫁接苗只表現出Tr和Gs下降,這可能是因為物種的差異性和脅迫物質的差異導致的,同時,我們試驗是在氣候箱中以水培方式進行的,這可能是造成Tr下降的因素之一。
植物葉綠素主要通過葉綠素熒光、光合電子傳遞和熱耗散3個途徑吸收耗散光能,這3種途徑間存在密切的此消彼長的相關性,葉綠素熒光變化可以反映光合作用和熱耗散的情況[22]。卜瑞方[23]研究表明,與對照相比,2 mmol·L-1處理2周后顯著降低了黃瓜幼苗葉片的NPQ和qP,添加Si顯著增加了CA脅迫下NPQ和qP。CA處理顯著降低了黃瓜幼苗葉片Fv/Fo,添加Si顯著增加了CA脅迫下PSII潛在活性。本試驗結果表明,0.5 mmol·L-1 CA處理后的黃瓜實生苗的Fv/Fm、Fv/Fo、qP和ΦPSII較對照顯著降低,表明CA脅迫會降低PSII的原初光能轉換效率,同時使PSII的潛在活性變弱,甚至導致光合系統受損[24]。與實生苗相比,嫁接增強了葉片的Fv/Fm和NPQ,提高了其光保護能力及原初光能轉換效率,從而降低了外源CA脅迫對光合系統的損傷程度。通過增加非輻射性熱耗散光系統能夠消耗PSII吸收的過剩光能,防止PSII反應中心由于吸收過剩光能導致的光氧化損傷。0.5 mmol·L-1CA處理降低了黃瓜實生苗葉片NPQ,使葉片PSII的潛在熱耗散能力下降,導致PSII在CA脅迫下更易遭受光抑制和光氧化;嫁接增加了CA處理下的黃瓜苗葉片的NPQ,提高了PSII的潛在熱耗散能力,在CA脅迫下,能夠減輕因PSII吸收過剩的光能而造成的光抑制和光破壞,從而起到保護PSII免受或減輕光抑制和光氧化的作用。0.5 mmol·L-1CA 處理顯著減小了黃瓜實生苗葉片的qP值,說明自毒脅迫減少了參與CO2固定的電子數量,阻礙葉片暗反應正常進行,最終導致光合效率低于嫁接苗[25]。與0.5 mmol·L-1 CA處理嫁接苗相比,同處理下黃瓜實生苗葉片中葉綠素含量低,使電子傳遞速率、光能轉化效率、和qP值等大幅度下降,導致光能向化學能轉化的過程受抑制,與前人研究一致,說明嫁接可以減輕自毒脅迫對黃瓜幼苗光合系統的損傷程度。
黃瓜實生苗與嫁接苗對肉桂酸引起的自毒脅迫表現出不同的生長和生理響應。CA脅迫后黃瓜實生苗的生長發育、生物量的積累及光合能力均受到顯著抑制,而嫁接苗受到的抑制作用較小或無顯著差異。葉綠素熒光成像分析結果表明,嫁接通過保護PSII免受或減輕CA脅迫引起的光抑制和光氧化,從而保證植株正常的生長發育。
參考文獻
[1] 楊建霞.日光溫室黃瓜連作障礙研究及防治對策[J].甘肅農業,2005(11):209.
[2] 孫光聞,陳日遠,劉厚誠.設施蔬菜連作障礙原因及防治措施[J].農業工程學報,2005,21(S2):184-188.
[3] INDERJI T,DUKE S O.Ecophysiological aspects of allelopathy[J].Planta,2003,217(4):529-539.
[4] YU J Q,YE S F,ZHANG M F,et al.Effects of root exudates and aqueous root extracts of cucumber(Cucumis sativus L.)and allelochemicals,on photosynthesis and antioxidant enzymes in cucumber[J].Biochemical Systematics and Ecology,2003,31(2):129-139.
[5] 呂衛光,張春蘭,袁飛,等.有機肥減輕連作黃瓜自毒作用的機制[J].上海農業學報,2002,18(2):52-56.
[6] DING J,SUN Y,CIAO C L,et al.Physiological basis of different allelopathic reactions of cucumber and figleaf gourd plants to cinnamic acid[J].Journal of Experimental Botany,2007,58(13):3765-3773.
[7] 吳鳳芝.外源酚酸對黃瓜自毒作用的生理生化機制研究[D].哈爾濱:東北農業大學,2002.
[8] 高旭,張古文,胡齊贊,等.自毒物質肉桂酸對豌豆幼苗生長及葉綠素熒光特性的影響[J].中國蔬菜,2013(8):44-49.
[9] 陳紹莉,周寶利,王茹華,等.嫁接對茄子根系分泌物中肉桂酸和香草醛的調節效應[J].應用生態學報,2008,19(11):2394-2399.
[10] 王茹華,曲光峰,張啟發,等.肉桂酸對番茄的化感效應研究[J].中國蔬菜,2014(3):29-32.
[11] 喻景權,杜堯舜.蔬菜設施栽培可持續發展中的連作障礙問題[J].沈陽農業大學學報,2000,31(1):124-126.
[12] 陳紹莉,周寶利,尹玉玲,等.茄子自毒物質脅迫下嫁接對其生長及土壤生化特性的影響[J].園藝學報,2010,37(6):906-914.
[13] 裴孝伯,李世誠,張福墁,等.溫室黃瓜葉面積計算及其與株高的相關性研究[J].中國農學通報,2005(8):80-82.
[14] YU J Q,SHOU S Y,QIAN Y R,et al.Autotoxic potential of cucurbit crops[J].Plant and Soil,2000,223(1/2):149-153.
[15] 王崇啟,劉淑梅,侯麗霞,等.蔬菜嫁接專題報道(四)茄子嫁接苗的應用現狀及改進方案[J].中國蔬菜,2013(7):15-18.
[16] BAZIRAMAKENGA R,SIMARD R R,LEROUX G D.Effects of benzoic and cinnamic acids on growth,mineral composition,and chlorophyll content of soybean[J].Journal of Chemical Ecology,1994,20(11):2821-2833.
[17] 朱春俠,姜偉,劉文,等.黃瓜嫁接育苗的研究進展[J].北方農業學報,2019,47(2):115-118.
[18] 肖春蘭.黃瓜和黑籽南瓜幼苗對肉桂酸響應差異的生理機制研究[D].杭州:浙江大學,2006.
[19] WEIR T L,PARK S W,VIVANCO J M.Biochemical and physiological mechanisms mediated by allelochemicals[J].Current Opinion in Plant Biology,2004,7(4):472-479.
[20] 劉俊祥,孫振元,巨關升,等.重金屬Cd2+對結縷草葉片光合特性的影響[J].核農學報,2009,23(6):1050-1053.
[21] 賈聯紅.嫁接西瓜化感效應的研究[D].四川雅安:四川農業大學,2012.
[22] 馬洪英,楊小玲,仝雅娜,等.4個水果型黃瓜葉片葉綠素熒光特性的比較研究[J].天津農業科學,2019,25(10):19-22.
[23] 卜瑞方.硅緩解黃瓜種子萌發和幼苗生長自毒作用的機理研究[D].蘭州:甘肅農業大學,2016.
[24] ALLEN D J,ORT D R.Impact of chilling temperatures onphotosynthesis in warm climate plants[J].Trends Plant Science,2001,1(6):36-42.
[25] 楊廣東,朱祝軍,計玉妹.不同光強和缺鎂脅迫對黃瓜葉片葉綠素熒光特性和活性氧產生的影響[J].植物營養與肥料學報,2002,8(1):115-118.
