斑馬魚cetp基因敲除模型的建立及其肝臟轉錄組學分析
2020-07-09 12:33:10馮冬萍商漢橋張虎軍楊夢溪任景怡
基礎醫學與臨床 2020年7期
馮冬萍,商漢橋,楊 航,4,張虎軍,4,張 停,楊夢溪,屠 強,4,任景怡*
(1.北京大學中日友好臨床醫學院, 北京 100029; 2.中日友好醫院 心內科,北京 100029;3.中國科學院 遺傳與發育生物學研究所, 北京 100101;4.中國科學院大學, 北京 100049)
動脈粥樣硬化性心血管疾病(atherosclerotic cardiovascular disease, ASCVD)是全球成人死亡的首要原因[1]。盡管強化他汀治療已使心血管病患者明顯獲益,但心血管疾病仍存在較大剩留風險[2-5],且與高密度脂蛋白膽固醇(high-density lipoprotein cholesterol, HDL-C)水平較低相關[6]。因此,升高HDL-C水平以降低ASCVD剩留風險的策略備受關注。目前正在研發的升高HDL-C的相關治療方法中最受關注的是膽固醇酯轉運蛋白(cholesteryl ester transfer protein, CETP)抑制劑[7]。CETP是主要由肝臟和脂肪組織合成的疏水性糖蛋白,在循環中CETP與HDL結合,將膽固醇酯從HDL轉運至富含載脂蛋白B(apolipoprotein B,ApoB)的極低密度脂蛋白膽固醇(very low-density lipoprotein cholesterol, VLDL-C)和低密度脂蛋白膽固醇(low-density lipoprotein cholesterol, LDL-C),同時將ApoB脂蛋白中的三酰甘油(triglyceride,TG)逆向轉運給HDL,加速了HDL的分解代謝,在膽固醇逆轉運過程中起重要作用[8]。因此,從理論上講,抑制CETP可能是升高HDL-C最有效的藥理作用[9]。
然而,CETP抑制劑相關臨床試驗結果存在不確定性[10-14],并且CETP在動脈粥樣硬化疾病中的作用機制尚有很多不明之處[15-16]。CETP抑制劑的動物實驗目前困難重重。首先,缺乏合適的疾病動物模型。常用的嚙齒類動物缺少cetp基因,與人類完全不同,無法直接用于CETP作用機制的研究。而將人類CETP引入小鼠,得出的結果又相互矛盾[17]。其次,在動脈粥樣硬化疾病的條件下,CETP對HDL的影響以及膽固醇逆轉運(reverse cholesterol transport,RCT)的作用仍未在體內被證實。迄今為止,尚無通過抑制CETP升高HDL水平詳細機制的相關報告。基于此,尋找更合理的疾病模型對CETP的研究至關重要。斑馬魚體型小,易于大量繁殖,胚胎和幼魚透明,全基因組測序完成,適用于很多種遺傳學、發育生物學、生理學等技術手段,是一個理想的脊椎動物模型。更重要的是,斑馬魚擁有保守且與人類高度相似的脂類代謝相關基因,特別是具有cetp基因,且脂蛋白分布與人類相似,對高脂食物敏感[18]。因此,斑馬魚非常適合脂代謝及動脈粥樣硬化研究,受到了廣泛關注。
本研究應用CRISPR/Cas9技術敲除斑馬魚cetp基因,構建cetp-/-模型,并且通過轉錄組測序探討cetp基因敲除后其他相關基因的變化及其相互作用,進而揭示CETP在動脈粥樣硬化疾病過程中的作用機制。
1 材料與方法
1.1 材料
主要試劑:Trizol 試劑(Invitrogen公司);2×Es Taq MasterMix(北京康為世紀生物科技公司);RNeasy Mini kit(Qiagen公司);反轉錄試劑盒(Bio-rad公司);無水乙醇(北京化工廠);3-氨基苯甲酸乙酯甲基磺酸鹽(3-aminobenzoic acid ethyl ester methanesulfonate,MS-222)(Sigma-Aldrich公司);T7核酸內切酶1(T7 endonuclease 1, T7E1)(BioLabs公司)。
1.2 方法
1.2.1 斑馬魚飼養:本實驗所用斑馬魚為野生型Tu品系,其飼養條件為28.5 ℃,pH 7.2~7.6,光照周期為10 h黑暗、14 h光照。斑馬魚繁殖后代方法:即選用健康性成熟的雌、雄斑馬魚,按雄∶雌=1∶1~2的數量置于產卵盒中,并用透明隔板分離雌、雄斑馬魚,次日移去隔板,收集胚胎。斑馬魚胚胎放入培養液中,于28 ℃恒溫培養箱中培養,受精后4~5 d(days post fertilization,dpf)轉到魚房并喂食草履蟲,10~14 dpf轉入北京愛生公司斑馬魚養殖系統中養殖喂食豐年蝦,斑馬魚胚胎發育時期按照Kimmel等的標準[19]確定。
1.2.2 斑馬魚cetp基因敲除:在NCBI GenBank獲得斑馬魚cetp的全長序列,應用CRISPR/Cas9靶位點設計網站SSC(http://crispr.dfci.harvard.edu/SSC/)設計sgRNA。在斑馬魚cetp第2個外顯子區域設計基因編輯靶位點,其堿基序列為5′-GGG ACCATCAACTATGGATA-3′。此外,設計正向引物487f(5′-CTGAAGTCATACAAGCCGC-3′)和反向引物494r(5′-CATTATGCATTAACTTGACT-3′)用于檢測靶位點是否發生基因編輯。體外合成sgRNA、Cas9核酸酶mRNA,將sgRNA、Cas9 mRNA及酚紅(指示劑)共孵育配制顯微注射液。收集斑馬魚一細胞期胚胎并注入上述顯微注射液。對照組為同一批未注射的斑馬魚胚胎。斑馬魚胚胎顯微注射操作參考John D.Mably等[20-21]的研究。將注射和未注射的斑馬魚胚胎均于28 ℃恒溫培養箱中培養,定期更換培養液。
收集顯微注射2 dpf胚胎和對照組胚胎各24枚,分別提取基因組DNA,并通過PCR反應及T7E1酶切確定注射胚胎是否發生基因編輯。酶切成功樣品的PCR產物送北京擎科新業生物技術有限公司測序。基因編輯成功的F0胚胎飼養至性成熟,并與野生型成魚雜交,所得胚胎提取基因組DNA后進行PCR擴增、T7E1酶切及測序,檢測靶位點突變情況。將突變胚胎飼養至成魚,通過剪尾提取F1基因組DNA并進行PCR擴增、T7E1酶切及測序,確定突變類型,從而獲得cetp基因敲除的雜合突變體,即cetp+/-。同基因型F1雌雄斑馬魚自交后代飼養至成魚,剪尾提取其基因組DNA并進行PCR擴增、T7E1酶切及測序,獲得cetp基因敲除純合突變體,即cetp-/-。
1.2.3 斑馬魚標本取樣:使用麻醉劑MS-222麻醉成年斑馬魚后,在顯微鏡下分離斑馬魚肝臟,用無RNA酶的PBS溶液沖洗后置于1 mL Triol中,于-80 ℃冰箱儲存。
1.2.4 RNA提取:取-80 ℃凍存的肝臟組織,于冰上勻漿。樣品于室溫下靜置5 min后加入200 μL三氯甲烷,充分搖晃15 s后,室溫靜置5 min;4 ℃,15 000×g離心15 min;取400 μL上層液體于新的1.5 mL離心管中,并加入400 μL 70%乙醇吹打混勻。后續實驗方法按照QIAGEN RNeasy Micro Kit使用說明進行。
1.2.5 轉錄組測序及分析:首先采用Nano Drop 2000微量分光光度計及Agilent 2100 Bioanalyzer,Agilent RNA 6000 Nano Kit對RNA進行純度、濃度與完整系數(RNA integrity number, RIN)檢測,RIN≥9.0、28S/18S<1.2的RNA用于建庫、測序。文庫構建及測序由北京安諾優達基因科技有限公司采用HiSeq Xten PE500測序平臺完成。通過轉錄組測序技術獲得基因表達譜變化,應用KEGG數據庫進行差異表達基因功能富集,分析其可能影響的生物學過程及信號通路。
1.2.6 實時熒光定量PCR:采用實時熒光定量PCR(real-time quantitative polymerase chain reaction,RT-qPCR)檢測斑馬魚肝臟cetp基因表達情況。Trizol法提取野生型和cetp-/-斑馬魚肝臟總RNA,以RNA為反轉錄模板,應用反轉錄試劑盒合成cDNA,再以cDNA為模板進行RT-qPCR,其中,正向引物為q1-1f:5′-AGCATGCCAAATATCCCAGT-3′,反向引物q1-1r:5′-AGAAGCAAACATATCGTGTAA-3′。 RT-qPCR反應程序為第1個循環 95 ℃ 10 min;第2~41個循環95 ℃ 15 s,60 ℃ 1 min;第42循環熔解曲線。每個反應進行3次生物學重復,以β-actin為內參對照,對照組為外對照,采用2-ΔΔCt法計算cetp基因在野生型斑馬魚和cetp-/-斑馬魚肝臟相對表達量,應用GraphPad prism8繪制柱狀圖。
1.3 統計學分析
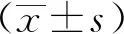
2 結果
2.1 應用CRISPR/Cas9技術構建cetp-/-斑馬魚模型
為獲得斑馬魚cetp基因敲除純合突變體,本研究應用CRISPR/Cas9技術對斑馬魚cetp基因第2個外顯子進行基因編輯(圖1A)。cetp基因敲除純合突變體遺傳學篩選流程(圖1B)如下:通過顯微注射技術將sgRNA、Cas9 mRNA及酚紅組成的顯微注射液注入野生型斑馬魚一細胞期胚胎,構建F0 founder;F0與野生型斑馬魚交配獲得F1,提取F1基因組DNA,通過PCR擴增及T7E1酶切實驗確定cetp+/-斑馬魚;根據孟德爾遺傳定律,同基因型F1雌、雄斑馬魚交配即可獲得cetp-/-斑馬魚。采用3對引物通過PCR擴增、T7E1酶切及瓊脂糖凝膠電泳篩選野生型、雜合體、純合體(圖1C),其中野生型cetp(泳道3、4、6、15、16, +/+表示)及cetp-/-純合突變體(泳道1、2、9, -/-表示)斑馬魚的PCR產物因不能被T7E1酶切,所以電泳結果呈現單1條帶(約390 bp),而cetp+/-雜合突變體(泳道5、7、8、10、11、12、13、14,+/-表示)因雜合雙鏈退火產生非配對DNA片段,T7E1可識別不完全配對DNA并進行酶切,酶切產物電泳后會形成2條帶,因而出現約290 bp和360 bp的2條帶。上述cetp基因敲除雜合體和純合體PCR產物進行TA克隆測序(圖1D),箭頭所示為基因編輯起始位點,產生了增加5個堿基TCCCT的移碼突變。由此,成功構建了cetp-/-斑馬魚模型。
2.2 驗證cetp-/-斑馬魚模型
通過RT-qPCR驗證野生型和cetp-/-斑馬魚的肝臟cetp轉錄水平。結果顯示,cetp-/-斑馬魚肝臟中cetp基因表達量是野生型斑馬魚表達量的約1/5,cetp-/-斑馬魚肝臟cetp基因表達量顯著降低(P<0.05)。
2.3 轉錄組測序數據分析
分別提取野生型和cetp-/-斑馬魚肝臟總RNA,利用轉錄組測序技術進行轉錄調控分析;根據測序數據分析上述樣品之間的相關性。結果顯示,野生型斑馬魚與cetp-/-斑馬魚肝臟樣品分為截然不同的兩類且組內相關性較好,該結果提示斑馬魚cetp基因敲除成功,與實驗預期相符,測序數據可進行后續轉錄組學分析(圖2)。

A.zebrafishcetpgene and knockout site; B.zebrafishcetp-/-model genetics screening process; C.T7E1 assay for detecting CRISPR/Cas9-mediatedcetpgene editing zebrafish; D.sequencing graphs of PCR product from zebrafish; +/+.wild type; +/-.heterozygote; -/-.homozygote
圖1 CRISPR/Cas9介導的斑馬魚cetp基因敲除
Fig 1 CRISPR/Cas9-mediated zebrafishcetpknockout

圖2 野生型和cetp-/-斑馬魚肝臟樣品的相關性分析
2.4 差異基因表達分析
從野生型斑馬魚和cetp-/-斑馬魚(測序)文庫中,通過P<0.05篩選表達量發生變化的差異表達基因(圖3)。圖中紅色代表顯著上調基因,藍色代表顯著下調基因。與野生型斑馬魚相比,cetp-/-斑馬魚共篩選到3 808個發生顯著變化的基因,其中cetp基因敲除后發生顯著上調的基因有1 918個,顯著下調的基因有1 890個。該結果提示cetp-/-斑馬魚與野生型斑馬魚肝臟基因表達具有顯著差異。

圖3 野生型和cetp-/-斑馬魚肝臟差異表達基因
2.5 KEGG分析
將篩選得到的差異表達基因于KEGG數據庫進行通路注釋,通過顯著性水平(P<0.05)篩選顯著差異的KEGG通路(表1)。其中,上調基因富集的通路包括溶酶體通路、自噬調節通路等;而下調基因富集的通路包括DNA復制通路、核糖體通路及三羧酸循環通路等。

表1 差異表達基因通路富集列表
3 討論
冠心病已成為全球死亡原因之首,嚴重威脅人類健康[1, 22],血脂異常是其主要危險因素之一[23]。盡管人類CETP突變后其功能缺失可升高血漿HDL-C、降低LDL-C水平、減少心血管事件[24],然而,CETP抑制劑臨床試驗顯示,抑制CETP而升高HDL-C濃度的治療策略并未顯著降低心血管剩留風險[10]。目前,CETP在動脈粥樣硬化中的作用機制仍未完全明確。因此,尋找理想動物模型深入研究CETP在動脈粥樣硬化的具體作用機制可為治療冠心病提供新的治療策略,具有重要臨床意義。斑馬魚以其獨特的生理優勢、清晰的遺傳背景和豐富的數據庫資源成為重要的脊椎動物模型。更為重要的是,斑馬魚具有cetp基因,其脂蛋白分布和人類相似,使其在脂質代謝及動脈粥樣硬化疾病的研究中獨具優勢[18, 25-27]。第三代基因組編輯技術CRISPR/Cas9是在sgRNA引導下使cas9蛋白定位到DNA靶序列上,對DNA雙鏈進行切割,實現基因編輯,具有設計簡單、操作方便、切割效率高等優勢[28]。
本研究通過CRISPR/Cas9和顯微注射技術敲除斑馬魚cetp基因,構建斑馬魚cetp-/-模型,并且分別對野生型和cetp-/-斑馬魚肝臟進行轉錄組學測序。經轉錄組分析,本研究共篩選到3 808個差異表達基因,并對其進行KEGG富集分析,結果顯示,敲除斑馬魚cetp基因后,abca1、abcg2a基因顯著上調;lipg基因顯著下調。Bochem等[29]研究顯示,因三磷酸腺苷結合盒轉運體A1(adenosine triphosphate-binding cassette transporter A1,ABCA1)突變導致ABCA1功能喪失后,頸動脈粥樣硬化增加,CVD風險增加,上調ABCA1成為治療CVD的有效治療策略。內皮脂酶是由lipg基因編碼產生的脂肪酶,主要參與脂蛋白代謝過程,是HDL代謝中的關鍵蛋白。Ishida等[30]將小鼠Lipg敲除后,血漿HDL顆粒直徑變大,HDL水平升高,HDL清除延遲;而將Lipg轉入小鼠體內后,血漿HDL水平降低。此外,人類中LIPG功能丟失,HDL水平顯著升高[31]。本研究中敲除斑馬魚cetp基因后abca1、abcg2a基因顯著上調;lipg基因顯著下調,可能cetp-/-斑馬魚的Hdl水平升高,具有抗動脈粥樣硬化作用。
此外,本研究還發現,與野生型斑馬魚相比,cetp-/-斑馬魚顯著上調基因主要富集到自噬相關通路,提示敲除cetp后斑馬魚自噬作用明顯增強。研究表明,自噬與炎性反應相互影響,二者均在動脈粥樣硬化中具有重要作用,適度自噬反應對機體具有保護作用,而過度自噬會引發細胞死亡及促進炎性因子分泌,可能加速動脈粥樣硬化斑塊不穩定[32-36]。新近的CANTOS研究首次證實抑制炎性反應可降低不良心血管事件,為動脈粥樣硬化的炎性反應假說提供直接證據[37]。因此,敲除cetp后斑馬魚的自噬作用顯著增加,促進炎性反應,可能具有促動脈粥樣硬化的作用。
綜上所述,盡管敲除斑馬魚cetp可導致血漿Hdl升高,改變脂質代謝,降低動脈粥樣硬化風險,但是敲除cetp后會激活自噬,增加炎性反應,具有促動脈粥樣硬化作用。本研究結果表明,CETP在動脈粥樣硬化中具有抗動脈粥樣硬化和促動脈粥樣硬化雙向調節作用,CETP在炎性反應中的具體作用機制仍需進一步研究。
