絲裂原活化蛋白激酶4通過激活AKT促進宮頸癌細胞增殖
2020-07-09 02:03:34賴麗梨段華英鄒爭志
激光生物學報 2020年3期
賴麗梨,段華英,鄒爭志
(1.廣州市增城區(qū)人民醫(yī)院婦產科, 廣州 511300; 2.廣州醫(yī)科大學附屬第二醫(yī)院婦產科, 廣州 510260;3.華南師范大學生物光子學研究院激光生命科學教育部重點實驗室, 廣州 510631; 4.華南師范大學生物光子學研究院, 廣東省激光生命科學重點實驗室, 廣州 510631)
絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)是一個能被不同的細胞外刺激激活的絲氨酸-蘇氨酸蛋白激酶家族,細胞外的因素包括細胞生長因子、神經遞質、激素、細胞應激及細胞黏附。MAPK信號通路是真核生物信號傳遞網絡中的重要途徑之一,在基因表達調控和細胞功能活動中發(fā)揮著關鍵作用。經典的MAPK家族可分為ERK、p38、JNK和ERK5 4個亞族。已知MAPK這4個家族成員在細胞的生長、分化、對環(huán)境的應激適應、炎癥反應等多種重要的細胞生理/病理過程中發(fā)揮著重要的作用。MAPK家族的過度激活在多種腫瘤中被發(fā)現(xiàn),MAPK的激活與腫瘤細胞的增殖、轉移和遷移有重要關系[1]。MAPK4是MAPK家族的一個非典型的成員,關于MAPK4的生物學功能以及與疾病的關系,目前報導的很少。有研究發(fā)現(xiàn),MAPK4能夠通過調節(jié)MAPK5的功能去影響細胞的增殖[2]。最近的研究發(fā)現(xiàn),MAPK4在前列腺癌中過表達,干擾MAPK4的表達顯著地抑制了癌細胞的增殖和錨定依賴的增長。同時,在小鼠體內的研究也發(fā)現(xiàn),抑制MAPK4活性能夠顯著地抑制移植瘤的生長[3]。通過研究MAPK4誘導腫瘤形成的分子機制,他們發(fā)現(xiàn)MAPK4通過激活mTOR信號通路去誘導腫瘤的發(fā)生發(fā)展。最近在骨髓瘤研究中,F(xiàn)eng等[4]發(fā)現(xiàn)MAPK4在腫瘤進展中起了重要作用,同時也發(fā)現(xiàn),CircRNA circ_0000190通過調控miR-767-5p抑制了MAPK4的表達,從而抑制骨髓瘤的發(fā)展。
經典的MAPK家族在宮頸癌中的研究已有一些報導,然而MAPK4在宮頸癌中的作用,目前尚沒有研究報導。在本研究中,我們發(fā)現(xiàn)MAPK4能夠激活宮頸癌細胞HeLa和SiHa中的蛋白激酶B(protein kinase B,AKT,又稱PKB)信號,進一步發(fā)現(xiàn)MAPK4的表達與宮頸癌細胞的增殖顯著相關。
1 材料與方法
1.1 試劑
DMSO購買于Sigma公司,MAPK4抗體購買于Abcam公司,p-AKT-Ser473、AKT和Actin抗體購買于Cell Signalling Technology公司,siRNA合成于上海吉瑪公司,LipofectamineTM3000、CCK8檢測試劑盒、胰蛋白酶消化液購買于Thermo Scientific公司。MAPK4過表達質粒由本課題組構建。
1.2 細胞培養(yǎng)
人宮頸癌細胞系HeLa和SiHa(購于中科院上海細胞庫),培養(yǎng)采用含10%胎牛血清(Gbico)的DMEM培養(yǎng)基,置于37 ℃、5%CO2、飽和濕度的細胞培養(yǎng)箱中常規(guī)培養(yǎng),經檢測培養(yǎng)箱無支原體污染[5]。
1.3 細胞增殖檢測
取對數(shù)生長期細胞,用胰酶消化后稀釋成每毫升5×104個細胞的懸液接種于96孔板中,每孔200 μL(即10 000個細胞)。在37 ℃、5%CO2培養(yǎng)箱中培養(yǎng),分別培養(yǎng)至24、48、72及96 h,按照CCK8檢測試劑盒說明書流程在上述時間點完成細胞增殖檢測。簡單的流程如下:試驗中止前4 h加入CCK8液20 μL,再培養(yǎng)4 h,在酶聯(lián)檢測儀上檢測450 nm 波長下每孔的吸光度OD(optical density)值,按下列公式求出細胞活力。細胞活力(cell viability)=(干擾或過表達MAPK4組平均OD值/對照組平均OD值)×100%[6]。
1.4 GSCA分析細胞系mapk4基因表達
通過在線軟件GSCA(http://bioinfo.life.hust.edu.cn/web/GSCALite/)[7],分析mapk4基因在已經檢測過的癌細胞系中mRNA的表達水平。根據(jù)mapk4基因的表達水平,選取了表達相對高的宮頸癌細胞HeLa和表達相對低的宮頸癌細胞SiHa。
1.5 細胞轉染
mapk4的siRNA干擾片段序列如下:5′-GGGU GAGCUGUUCAAGUUCTT-3′。對照序列(negative control,NC):5′-UCCGUUUCGGUCCACAUUC-3′。轉染時根據(jù)LipofectamineTM3000說明書,用適量無血清培養(yǎng)液將siRNA(終濃度100 nmol/L)或MAPK4過表達質粒(6孔板中每孔加入2 μg)與LipofectamineTM3000混勻,然后加入到細胞中,8 h后換為含10%胎牛血清培養(yǎng)液繼續(xù)培養(yǎng),在轉染后48 h內完成免疫印跡試驗,在轉染后96 h內完成細胞增殖檢測[8]。
1.6 免疫印跡
將處理過的細胞用胰酶消化收集,用預冷的PBS洗3遍,加入適量裂解液(50 mmol/L Tris-HCl pH 8.0、150 mmol/L NaCl、1%Triton X-100、1 mmol/L Na3VO4、100 mmol/L PMSF)在冰上裂解30 min,用移液槍反復吹打40次,避免產生氣泡。12 000 r/min離心15 min,收集上清蛋白液。采用10%~12%SDS-聚丙烯酰胺凝膠電泳,然后轉移到聚偏二氟乙烯(polyvinylidene fluoride, PVDF)膜上,轉膜完成后,用5%奶粉液或5%的牛血清白蛋白(bovine serum albumin,BSA)在室溫封閉1 h,之后分別在4 ℃孵育一抗12 h,在室溫孵育二抗1 h。ECL(electrochemiluminescence)底物顯色,A、B液以1∶1的體積比例混合,用濾紙將膜表面液體吸干,加ECL底物顯色液,放入暗盒中并壓片,5 s~5 min后顯影、定影[9]。
1.7 統(tǒng)計學分析
統(tǒng)計學方法數(shù)據(jù)均采用SPSS16.0軟件進行統(tǒng)計學分析,所有試驗重復3次,均為獨立試驗,計量資料采用均數(shù)±標準差表示,數(shù)據(jù)的比較分析采取student’t檢驗分析,P<0.05為差異有統(tǒng)計學意義。
2 結果與分析
2.1 mapk4在癌細胞系中的差異表達
為了評估m(xù)apk4在腫瘤細胞系中的表達, 通過GSCA在線軟件,在12個組織類型共65種腫瘤細胞系中,我們分析了mapk4的mRNA水平。
如圖1a所示,mapk4在肺癌細胞SCLC-21H中表達最高,其次是宮頸癌細胞HeLa。本研究選取了mapk4高表達的HeLa細胞和mapk4表達相對低的宮頸癌細胞SiHa作為研究的細胞模型。首先,通過RT-PCR試驗,我們在這兩個宮頸癌細胞中驗證了mapk4的表達,如圖1b所示,發(fā)現(xiàn)相對于HeLa細胞,SiHa細胞中mapk4的mRNA表達顯著降低。接下來,我們通過Western blot試驗,在這兩個細胞系中進一步驗證MAPK4蛋白的表達水平。如圖1c所示,與mapk4的mRNA表達結果一致,MAPK4的蛋白表達在HeLa細胞中顯著高于SiHa細胞。
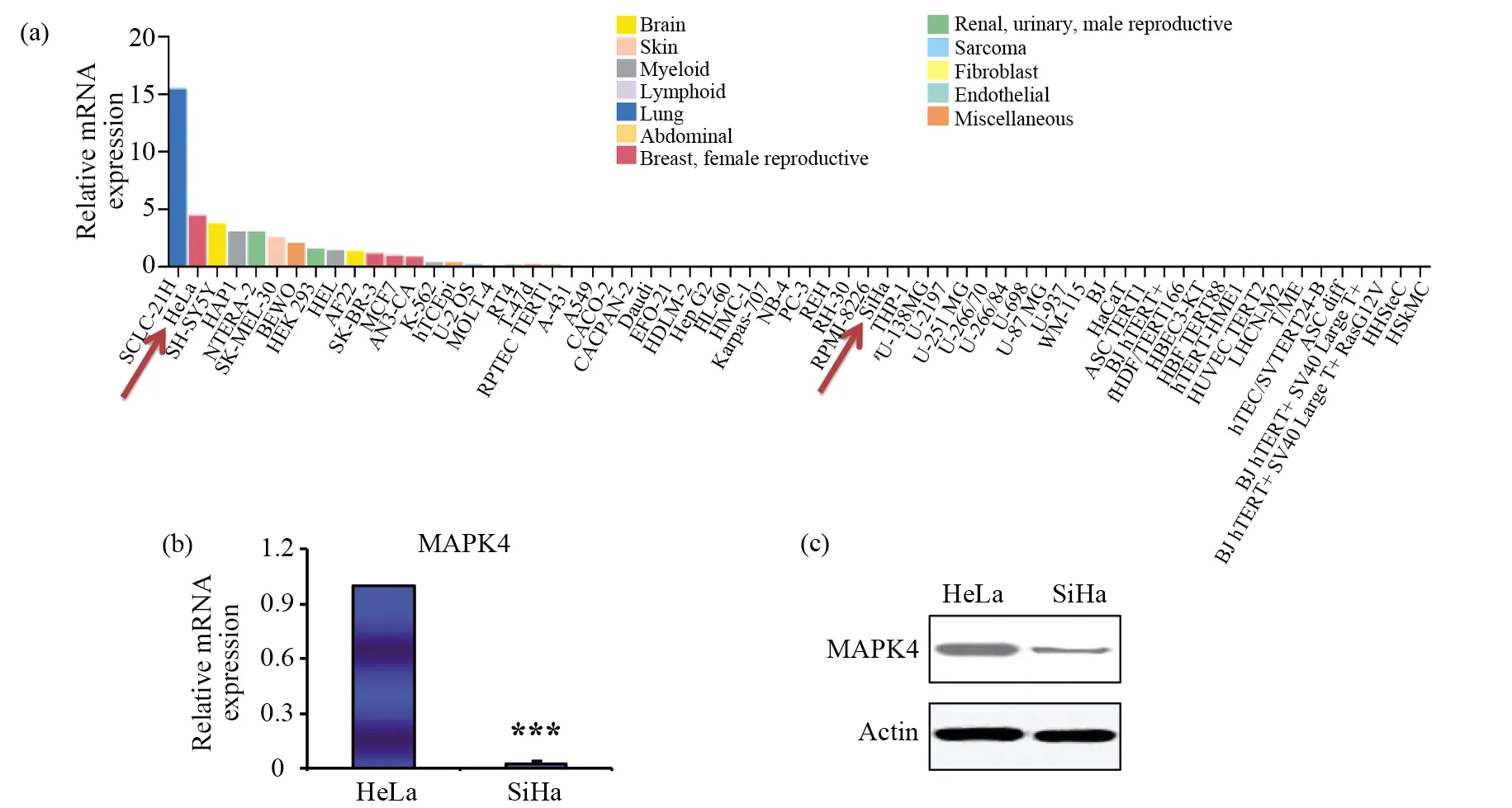
圖1 mapk4在癌細胞系中的差異表達Fig.1 Expression of mapk4 mRNA in cancer cell lines(a)通過在線軟件GSCA(http://bioinfo.life.hust.edu.cn/web/GSCALite/)[8]分析mapk4在已經檢測過的癌細胞系中mRNA的表達水平;(b)通過RT-PCR檢測HeLa和SiHa細胞中mapk4的mRNA水平。*** P<0.001,顯示有統(tǒng)計學顯著差異;(c)通過Western blot檢測HeLa和SiHa細胞中MAPK4的蛋白水平。Actin作為內參蛋白(a)The mapk4 mRNA levels of cancer cell lines are obtained from GSCA database(http://bioinfo.life.hust.edu.cn/web/GSCALite/);(b) mapk4 mRNA levels were detected by RT-PCR in HeLa and SiHa cells. *** P<0.001 indicates significant difference;(c)MAPK4 protein levels were detected by Western blot in HeLa and SiHa cells. Actin serving as loading control
2.2 MAPK4表達影響宮頸癌細胞增殖能力
MAPK家族蛋白在促進細胞增殖過程中起了重要作用。為探索MAPK4是否與宮頸癌細胞的增殖能力相關,本研究選取了MAPK4表達相對高的HeLa細胞,通過轉染mapk4的siRNA完成了MAPK4敲低試驗,并通過CCK8(cell counting kit-8)試驗檢測了細胞的增殖能力。如圖2a所示,在轉染了mapk4的siRNA后,MAPK4的蛋白表達顯著降低。細胞活力試驗也表明,干擾MAPK4后,細胞的增殖能力顯著降低,具體見圖2b。同時,我們選取了MAPK4相對表達低的SiHa細胞,在細胞中轉染MAPK4的過表達質粒,然后通過CCK8試驗檢測了細胞的增殖能力。如圖2c所示,在轉染了MAPK4的過表達質粒后,MAPK4的蛋白表達顯著升高。細胞活力試驗也表明,高表達MAPK4蛋白后,細胞的增殖能力顯著增強(圖2d)。以上結果表明,抑制MAPK4表達能夠抑制細胞的增殖能力,而促進MAPK4蛋白表達能夠增強宮頸癌細胞的增殖能力。這些都說明在宮頸癌中MAPK4可能和腫瘤細胞的增殖能力有關。
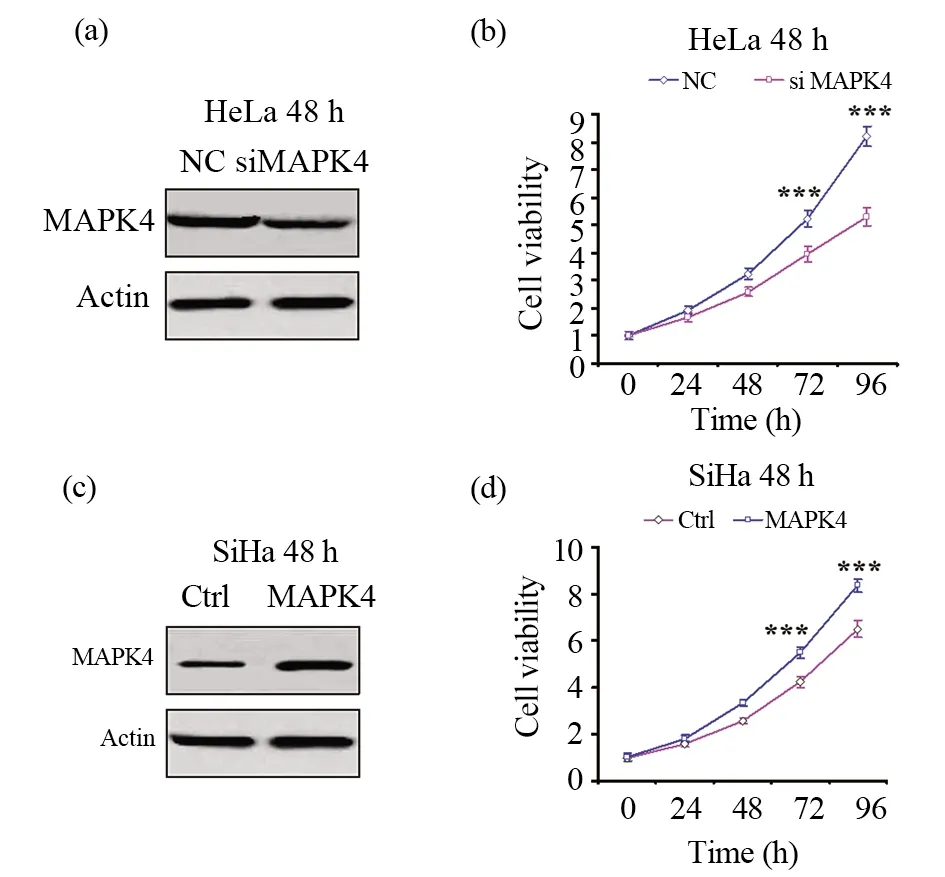
圖2 MAPK4表達影響宮頸癌細胞增殖能力Fig.2 MAPK4 expression is associated with cell proliferation in cervical cancer cell lines(a)在HeLa細胞中轉染mapk4的siRNA 48 h后,通過Western blot檢測MAPK4的表達,Actin作為內參蛋白;(b)在HeLa細胞中轉染mapk4的siRNA 48 h后,通過CCK8試驗檢測圖中所示時間點的細胞增殖能力;(c) 在SiHa細胞中轉染MAPK4過表達質粒48 h后,通過Western blot檢測MAPK4的表達,Actin作為內參蛋白;(d)在SiHa細胞中轉染MAPK4過表達質粒48 h后,通過CCK8試驗檢測圖中所示時間點的細胞增殖能力。NC:陰性對照;Ctrl:對照。*** P<0.001,顯示有統(tǒng)計學顯著差異(a)HeLa cells were transfected MAPK4 siRNA, after 48 h, MAPK4 were detected by Western blot,Actin serving as loading control;(b) HeLa cells were transfected MAPK4 siRNA, after 48 h, cell viability was detected by CCK8 assay at the time points shown in the figure;(c)SiHa cells were transfected MAPK4-overexpressed plasmid. After 48 h, MAPK4 was detected by Western blot. Actin serving as loading control;(d)SiHa cells were transfected MAPK4-overexpressed plasmid, after 48 h, cell viability was detected by CCK8 assay at the time points shown in the figure. NC:negativecontrol;Ctrl:Control. *** P<0.001indicates significant difference
2.3 在宮頸癌細胞中MAPK4促進了AKT的活性
以上的試驗研究結果表明,MAPK4促進了細胞的增殖能力,然而其下游的分子機制尚不清楚,因此,接下來我們探索了MAPK4下游與細胞增殖可能相關的蛋白。在肺癌細胞中的研究發(fā)現(xiàn)MAPK4激活了AKT信號,因此,在本研究中我們探討了MAPK4在宮頸癌細胞中是否也能激活AKT。我們同樣地選取了MAPK4表達相對高的HeLa細胞,通過轉染mapk4的siRNA完成了MAPK4敲低試驗。接下來,通過Western blot試驗檢測了AKT的活化形式磷酸化AKT(p-AKT-Ser473)的表達。如圖3a所示,在轉染了mapk4的siRNA后,MAPK4的蛋白表達顯著降低,同時p-AKT-Ser473的表達水平也顯著降低。同時,我們選取了MAPK4表達相對低的SiHa細胞,通過轉染MAPK4的過表達質粒,檢測了p-AKT-Ser473的表達水平。如圖3b所示,在轉染了MAPK4的過表達質粒后,MAPK4的蛋白表達顯著升高,同時p-AKT-Ser473的表達水平也顯著上升。以上結果表明,抑制MAPK4表達能夠抑制AKT的活性,而促進MAPK4表達能夠增強AKT的活性。這說明在宮頸癌中MAPK4可能通過激活AKT增強了腫瘤細胞的增殖能力。

圖3 在宮頸癌細胞中MAPK4促進了AKT的活性Fig.3 MAPK4 activates AKT in cervical cancer cells(a)和 (b)在宮頸癌細胞HeLa和SiHa中,分別轉染mapk4的siRNA和MAPK4過表達質粒,48 h后通過Western blot檢測MAPK4和p-AKT-Ser473的表達。AKT作為p-AKT-Ser473的內參蛋白,Actin作為總的內參蛋白(a) and (b) HeLa and SiHa cells were transfected mapk4 siRNA and MAPK4-overexpressed plasmid respectively, after 48 h, MAPK4 and p-AKT were detected by Western blot. AKT serving as loading control for p-AKT, Actin serving as loading control for total protein
3 討論
在全世界宮頸癌是女性第三常見的癌癥[10]。隨著篩選方法的不斷改進以及發(fā)達國家的疫苗接種計劃,發(fā)達國家與資源匱乏的發(fā)展中國家的婦女在宮頸癌發(fā)病率方面差異越來越顯著[11]。目前,大于85%的宮頸癌死亡發(fā)生在中低收入人群的國家。已知宮頸癌的發(fā)生與人乳頭瘤病毒(human papilloma virus,HPV)感染顯著相關,然而在沒有感染HPV的人群中也有一部分人會發(fā)展為宮頸癌[12],這可能和遺傳以及病毒之外的環(huán)境因素有關。現(xiàn)已有研究表明,一些原癌基因的突變能夠引起宮頸癌的發(fā)生,因而靶向抑制這些癌基因對宮頸癌的治療有潛在的價值[13]。在本研究中,我們發(fā)現(xiàn)MAPK4促進了宮頸癌細胞的增殖,因而MAPK4可以作為宮頸癌治療的潛在靶點。但本研究我們僅用到了2個宮頸癌細胞系驗證MAPK4促進宮頸癌細胞增殖,缺乏動物試驗的結果。因此,下一步需要構建動物模型,在體內驗證MAPK4促進了宮頸癌的增殖,同時,也需要在臨床組織標本中檢測MAPK4的表達是否與病人的預后相關。
在大多數(shù)人類腫瘤中,AKT蛋白是最常見的、活化的癌基因產物。AKT活化的主要機制包括2個主要途徑:一是通過腫瘤抑制基因pten突變,從而使PTEN失活,PTEN是AKT的一個抑制蛋白,因此PTEN突變的失活會導致AKT的過度激活;二是PI3K酶活性亞基的活化突變,pi3k是AKT蛋白的直接上游基因,PI3K能夠直接磷酸化激活AKT,因此PI3K的突變激活能夠促進AKT的過度激活[14,15]。最近越來越多的證據(jù)表明,除了上述兩條途徑外還有一些蛋白的表達和活性失調也影響了AKT的活性。比如ras基因突變導致Ras蛋白的過度活化也直接影響了AKT的活性,另外,akt基因自身的突變也是導致其過度活化的原因,如發(fā)生在AKT蛋白的pH功能區(qū)E17位的突變[16]。akt基因的擴增導致AKT蛋白的過度表達也是AKT過度活化的原因之一。在本研究中,我們發(fā)現(xiàn)MAPK4過表達能夠激活AKT的活性,然而具體的分子機制仍然不清楚。MAPK4是否直接與AKT相互作用,從而磷酸化AKT?還是通過激活一個中間分子間接激活AKT?這其中的分子機制還有待進一步研究。最近在肺癌的研究報道中發(fā)現(xiàn)MAPK4激活了AKT的活性,然而關于MAPK如何激活AKT活性的具體機制并未闡明。AKT促進細胞的增殖主要涉及以下幾種機制:1)抑制細胞周期抑制蛋白p27表達,p27蛋白能夠與細胞周期相關蛋白激酶(cyclin-dependent kinases,CDK)結合,從而抑制CDK的活性,進而阻止細胞周期的進展[17];2)促進細胞糖代謝,為細胞分裂增殖提供營養(yǎng)[18];3)抑制GSK3β的活性,GSK3β能夠促進細胞周期蛋白cyclinD1的降解,從而使細胞周期從G1向S期轉化受到阻止,而AKT能夠抑制GSK3β的活性,因此促進了cyclinD1的表達,從而加速細胞周期從G1向S期轉化[19]。本研究中我們發(fā)現(xiàn)MAPK4激活了AKT,然而并沒有檢測AKT下游促進細胞增殖的信號。因此,在接下來的研究中,需要進一步檢測AKT下游的信號是否受MAPK4的影響。
總之,本研究發(fā)現(xiàn)MAPK4促進宮頸癌細胞的增殖,這表明在宮頸癌中靶向抑制MAPK4能夠抑制宮頸癌的進展,MAPK4可以作為宮頸癌治療的潛在靶點。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
