鞣花酸對變異鏈球菌的體外抑菌作用及機制研究
2020-07-09 10:47:17閆莉周曉英
中國藥房 2020年5期
關鍵詞:機制
閆莉 周曉英

中圖分類號 R965 文獻標志碼 A 文章編號 1001-0408(2020)05-0607-05
DOI 10.6039/j.issn.1001-0408.2020.05.20
摘 要 目的:研究鞣花酸對變異鏈球菌的體外抑菌作用及可能機制,為其防治齲病的應用提供實驗依據。方法:以復方氯己定含漱液為陽性對照、5%二甲基亞砜(DMSO)溶液為陰性對照,采用打孔法抑菌試驗測定抑菌圈直徑考察50、25、12.5、6.25、3.125、1.562 5 mg/mL鞣花酸對變異鏈球菌的抑菌效果,并采用微量稀釋法測定其對變異鏈球菌的最低抑菌濃度(MIC)和最低殺菌濃度(MBC)。以5%DMSO溶液為陰性對照,采用結晶紫染色法測定1/8MIC、1/4MIC、1/2MIC、MIC鞣花酸對變異鏈球菌生物膜形成的影響,并在熒光染色后通過激光掃描共聚焦顯微鏡觀察1/2MIC鞣花酸作用后生物膜的結構變化;分別采用苯酚硫酸法和還原型輔酶Ⅰ氧化法考察1/8MIC、1/4MIC、1/2MIC、MIC鞣花酸對變異鏈球菌細胞外多糖(EPS)的抑制率和細胞外基質中乳酸脫氫酶(LDH)活力的影響。結果:12.5~50 mg/mL鞣花酸對變異鏈球菌產生了直徑均大于15 mm的抑菌圈,50 mg/mL鞣花酸作用下抑菌圈直徑與復方氯己定含漱液相當。鞣花酸對變異鏈球菌的MIC、MBC分別為12.5、25 mg/mL。1/8MIC~MIC鞣花酸作用后細菌生物膜的存活率較陰性對照顯著降低(P<0.01),且呈劑量依賴趨勢,MIC鞣花酸作用后細菌生物膜的存活率僅為(16.41±1.346)%;熒光染色后顯微鏡下可見,1/2MIC鞣花酸作用后細菌生物膜結構被破壞;1/8MIC~MIC鞣花酸作用后細菌水不溶性EPS和水溶性EPS的抑制率較陰性對照均顯著升高(P<0.01),1/4MIC~MIC鞣花酸作用后細胞外基質中LDH活力較陰性對照均顯著升高(P<0.01),且呈劑量依賴趨勢。結論:鞣花酸對變異鏈球菌的生長具有一定的抑制作用;其機制可能與抑制EPS產生、降低細菌的黏附、破壞細菌細胞膜有關。
關鍵詞 鞣花酸;變異鏈球菌;細菌生物膜;體外;抑菌作用;機制
Study on in vitro Anti-bacterial Activity and Mechanism of Ellagic Acid on Streptococcus mutans
YAN Li1,ZHOU Xiaoying2(1.Central Laboratory, Xinjiang Medical University, Urumqi 830011, China;2.School of Pharmacy, Xinjiang Medical University, Urumqi 830011, China)
ABSTRACT? ?OBJECTIVE: To study the in vitro anti-bacterial activity and potential mechanism of ellagic acid on Streptococcus mutans, and to provide evidence for its prevention and treatment of dental caries. METHODS: Using Compound chlorhexidine gargle as positive control, 5%DMSO as negative control, bacteriostasis experiment was conducted by the method of drilling hole, and bacteriostatic effects of 50, 25, 12.5, 6.25, 3.125, 1.562 5 mg/mL ellagic acid on S. mutans was preliminarily determined by measuring the diameter of bacteriostatic ring. The minimal inhibitory concentration (MIC) and minimal bactericidal concentration (MBC) of ellagic acid on S. mutans were determined by microdilution method. Using 5% DMSO as negative control, the effects of 1/8 MIC, 1/4 MIC, 1/2 MIC and MIC ellagic acid on the formation of S. mutans biomembrane was determined by crystal violet staining. The changes of the biomembrane structure under the action of 1/2 MIC ellagic acid were observed by microscopy after fluorescence staining. Phenol sulfuric acid method and reducing coenzymeⅠoxidation method were used to determine inhibitory effects of 1/8MIC, 1/4MIC, 1/2MIC, MIC ellagic acid on S. mutans on extracellular polysaccharide (EPS) as well as effect on the activity of lactate dehydrogenase (LDH) in extracellular matrix. RESULTS: Ellagic acid with concentration of 12.5~50 mg/mL produced an inhibitory ring on S. mutans with diameter greater than 15 mm. Under the action of 50 mg/mL ellagic acid, the diameter of bacteriostatic ring was the same as that of Compound chlorhexidine gargle. MIC and MBC of ellagic acid to S. mutans were 12.5 mg/mL and 25 mg/mL. The survival rate of bacterial biomembrane after 1/8MIC-MIC ellagic acid treatment was significantly lower than that of the negative control (P<0.01), and had a certain dose-response trend. After MIC ellagic acid treatment, the survival rate of bacterial biomembrane was (16.41±1.346)%. After fluorescence staining, the structure of bacterial biomembrane was destroyed by 1/2 MIC ellagic acid. After treated with 1/8MIC-MIC ellagic acid, its inhibitory rates on water-soluble EPS and water-insoluble EPS were increased significantly, compared with negative control (P<0.01). After treated with 1/4MIC-MIC ellagic acid, the activity of LDH in the extracellular matrix of bacteria increased significantly, compared with negative control (P<0.01), in dose-effect dependent trend. CONCLUSIONS: Ellagic acid can inhibit the growth of S. mutans, the mechanism of which may be associated with inhibiting EPS production, reducing bacterial adhesion, destroying bacterial cell membrane.
KEYWORDS? ?Ellagic acid; Streptococcus mutans; Biomem- brane; in vitro; Anti-bacterial activity; Mechanism
齲病是一種口腔內多因素疾病,與心血管疾病、癌癥一起被列為人類三大重點防治的慢性非傳染性疾病[1]。雖然齲病的病程發展較為緩慢,但是其發病率高、流行區域廣泛,不容忽視。細菌的存在是齲病發生的基礎,而牙菌斑生物膜是齲病發生所依賴的微環境[2]。變異鏈球菌是目前公認的主要致齲菌群,常黏附于牙面形成生物膜而導致齲病[3]。糖代謝是變異鏈球菌致齲的重要途徑:變異鏈球菌以蔗糖為底物,可以合成細菌胞外多糖(EPS),而合成EPS和形成生物膜的能力是其經典毒力表型,這些毒力因子在變異鏈球菌致病過程中發揮著重要作用[4]。
目前,抗生素、氟化物以及免疫防齲等的使用已被證明能有效地抑制口腔內的致齲菌,但這些方法存在會產生耐藥性或致使口腔菌群失調等問題[5]。而天然產物因其毒副作用小的優勢,逐漸成為防齲藥物研究開發的熱點[6]。鞣花酸作為一種多酚二內酯成分,是沒食子酸的二聚衍生物,存在于許多水果(如覆盆子、石榴等)及藥用植物(如五倍子、地榆等)中[7]。鞣花酸因具有多種生物活性(如抗氧化、抗癌、抗突變、抑制潰瘍、抑制人體免疫缺陷病毒等)而受到廣泛關注[8-9]。近年來,許多多酚類化合物(如茶多酚、厚樸酚等)被證實具有廣譜的抑菌效果,且對口腔致齲菌有良好的抑制作用[10]。鞣花酸作為一種天然的多酚類成分,國內外對其抑菌作用,特別是其對口腔致齲菌的抑制作用及機制的研究報道相對較少。因此,本研究以鞣花酸為研究對象,通過考察對口腔主要致齲菌——變異鏈球菌的生長、細菌生物膜形成等毒力因子的影響,探索其在防治齲病方面的開發應用潛力。
1 材料
1.1 儀器
C2型激光掃描共聚焦顯微鏡(日本Nikon公司);311型二氧化碳培養箱(美國Thermo公司);752型紫外分光光度計(上海菁華科技儀器有限公司);4K15型冷凍離心機(美國Sigma公司);LDZX-75KBS型高壓蒸汽滅菌鍋(上海申安醫療器械有限公司);SMP500型酶標儀(德國Spectro Max公司)。
1.2 藥品與試劑
復方氯己定含漱液(江蘇晨牌邦德藥業有限公司,批號:201810232,規格:500 mL,含葡萄糖酸氯己定0.6 g、甲硝唑0.1 g);鞣花酸對照品(中國食品藥品檢定研究院,批號:111959-201802,純度:≥98%);腦心浸肉湯(BHI)培養基(批號:1226L031)、刃天青染色劑(批號:512C031)均購自北京索萊寶科技有限公司;活/死細菌染色試劑盒(貨號:L13152)、磷酸鹽緩沖液 (PBS,批號:NXL0739)均購自賽默飛世爾生物化學制品(北京)有限公司;乳酸脫氫酶(LDH)試劑盒(中國南京建成生物工程公司,批號:20181025);其余試劑均為分析純,水為滅菌蒸餾水。
1.3 菌種
變異鏈球菌標準菌株(ATCC 700610)由新疆醫科大學中心實驗室提供。
2 方法
2.1 菌懸液的制備
取常規復蘇的變異鏈球菌標準菌株接種于BHI液體培養基中,于37 ℃培養箱中培養24 h后,取菌液在BHI板上劃線培養24 h,然后挑取單菌落在BHI液體培養基中以同樣條件純培養。待細菌進入對數生長期后,用紫外分光光度計調整菌液濃度約107 CFU/mL[在630 nm波長處的吸光度(OD)為0.2],備用。
2.2 抑菌圈的測定
采用打孔法抑菌試驗進行抑菌圈測定。將鞣花酸對照品用5%二甲基亞砜(DMSO)溶液溶解后制成質量濃度為50 mg/mL的母液;再用BHI液體培養基將母液按二倍稀釋法稀釋成鞣花酸質量濃度分別為25、12.5、6.25、3.125、1.562 5 mg/mL的藥液。取100 μL菌懸液(107 CFU/mL,下同)均勻涂布在BHI平板上,再以打孔器打孔。設置鞣花酸不同質量濃度組(1.562 6~50? ? mg/mL)、陰性對照組(5%DMSO)和陽性對照組(復方氯己定含漱液),每組重復設置3個孔,每孔中加入20 μL相應樣品溶液,然后于37 ℃培養箱中培養24 h。取出后,以十字交叉法測定抑菌圈直徑,取平均值。結果判定按《藥理實驗方法學》標準[11]:抑菌圈直徑<10 mm為無抑菌活性,10 mm為輕度敏感,11~15 mm為中度敏感,16~20 mm為高度敏感。
2.3 最小抑菌濃度的測定
使用96孔板,按100 μL/孔加入菌懸液,設置鞣花酸不同質量濃度組(50、25、12.5、6.25、3.125、1.562 5 mg/mL)、陰性對照組(5%DMSO溶液)和陽性對照組(復方氯己定含漱液),每組重復設置3個孔。每孔中加入100 μL相應樣品溶液,然后37 ℃培養箱中培養24 h。每孔中加入刃天青染色劑(1 mg/mL)20 μL,4 h后觀察,由藍色變為粉色的藥物濃度即為最小抑菌濃度(MIC)。
2.4 最小殺菌濃度的測定
根據MIC值測定結果,選擇藥液濃度≥MIC的各孔菌懸液,分別劃線于BHI平板上,將平板置于37 ℃培養箱中培養24 h,觀察無可見菌落生長的最低濃度即為最小殺菌濃度(MBC)。
2.5 變異鏈球菌細菌生物膜形成情況考察
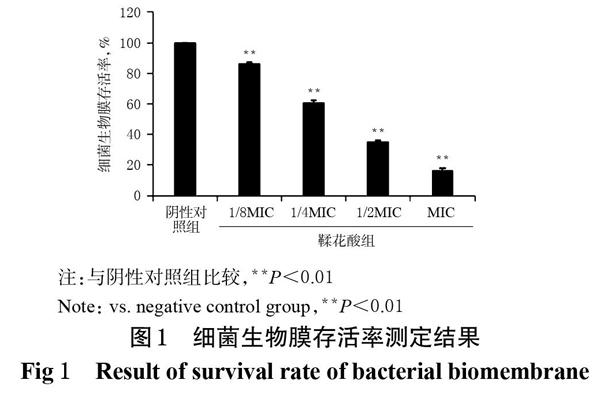
采用結晶紫染色法進行測定。使用96孔板,按100 μL/孔加入菌懸液,設置鞣花酸不同質量濃度組(1/8MIC、1/4MIC、1/2MIC、MIC)、陰性對照組(5%DMSO溶液),每組重復設置3個孔,每孔加入100 μL相應樣品溶液,然后置于37 ℃培養箱中培養24 h后,吸棄上清液。沉淀用無菌PBS輕輕洗滌2次,加入100 μL 0.1%結晶紫,對形成的生物膜染色15 min。棄去染料,用無菌PBS沖洗2次,干燥后每孔加入100 μL 95%乙醇溶液,溶解已被染色的生物膜上的結晶紫。采用酶標儀于540 nm波長處測定其OD值,并計算細菌生物膜存活率:細菌生物膜存活率(%)=1-[(1-OD給藥組/OD陰性對照組)×100%]。
2.6 變異鏈球菌細菌生物膜結構的觀察
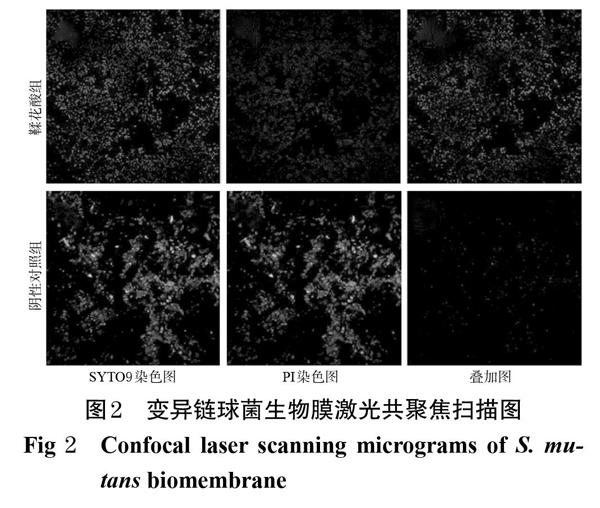
采用激光掃描共聚焦顯微鏡進行觀察。按500? ? ? ?μL/皿的量將菌懸液加入激光掃描共聚焦專用培養皿中,設置鞣花酸組(1/2MIC)和陰性對照組(5%DMSO溶液),每組設置3個平行皿,每皿加入500 μL相應樣品溶液,置于37 ℃培養箱中培養24 h后,吸棄上清液,以無菌PBS洗去浮菌,用2%甲基纖維素固定后,加入制備好的活/死細菌熒光染料20 μL[按說明書將試劑盒中熒光染色劑SYTO9和碘化丙啶(PI)按照 1 ∶ 1(V/V)的比例混合溶于適當體積的無菌水中],室溫下避光孵育20 min,然后立即在激光掃描共聚焦顯微鏡下觀察。活菌被SYTO9染色后發出綠色熒光,死菌被PI染色后發出紅色熒光,SYTO9和PI染色疊加圖中黃色表示活菌和死菌重疊處。
2.7 變異鏈球菌中EPS生成情況考察
采用苯酚硫酸法進行測定。按500 μL/管的量將菌懸液加入無菌離心管中,以5%DMSO溶液為陰性對照,考察不同質量濃度(1/8MIC、1/4MIC、1/2MIC、MIC)鞣花酸對變異鏈球菌中EPS生成的影響,每個樣品設置3個平行管,每管中加入5 mL相應樣品溶液,然后置于37 ℃培養箱中培養16 h。在4 ℃條件下以5 000 r/min離心20 min(下同),收集上清液;用無菌蒸餾水重懸沉淀并再次離心,收集上清液,合并2次上清液(含有水溶性EPS)。然后加入0.1 mol/L NaOH溶液5 mL重懸沉淀,離心2次,收集上清液(含有水不溶性EPS)。以0.22 μm的濾膜分別過濾以上2種上清液,各取適量分別加入3倍量冷卻的95%乙醇溶液,4 ℃過夜以沉淀EPS。次日再次離心,棄水相,沉淀物分別加入5 mL蒸餾水和5 mL 0.1 mol/L NaOH 溶液溶解。取上述含2種EPS的溶液各1 mL,分別加入5%冰苯酚1 mL,搖勻,迅速加入濃硫酸5 mL,搖勻,放置5 min后,放入沸水浴中反應15 min后,冷卻到室溫。采用紫外分光光度儀于490 nm波長處測定其OD值,并分別計算水溶性EPS和水不溶性EPS的抑制率:EPS抑制率(%)=(1-OD給藥組/OD陰性對照組)×100%。
2.8 變異鏈球菌細胞外基質中LDH活力的測定
采用還原型輔酶Ⅰ氧化法進行測定。使用96孔板,按100 μL/孔加入菌懸液和等量的BHI液體培養基后,置于37 ℃培養箱中培養24 h。棄去上清液,沉淀用無菌PBS緩沖液清洗2次。設置鞣花酸不同質量濃度組(1/8MIC、1/4MIC、1/2MIC、MIC)和陰性對照組(5%DMSO溶液),每組設置3個平行孔,在清洗過的培養孔(含有沉淀)中加入200 μL相應樣品溶液后,置于37 ℃培養箱中培養24 h。離心,收集上清液,按照LDH試劑盒說明書操作測定LDH活力。
2.9 統計學方法
采用SPSS 12.0軟件對試驗數據進行統計分析。計量資料以x±s表示,采用單因素方差分析的Dunnett(雙側)檢驗進行組間比較。P<0.05為差異有統計學意義。
3 結果
3.1 抑菌圈測定結果
結果顯示,5%DMSO作為溶劑對變異鏈球菌的生長無明顯變化。12.5~50 mg/mL鞣花酸對變異鏈球菌產生了抑菌圈,且抑菌圈的直徑隨著藥物質量濃度的增加而增大,其中50 mg/mL鞣花酸對變異鏈球菌抑菌圈的大小與復方氯己定含漱液相近,對變異鏈球菌均為高度敏感。抑菌圈直徑測定結果見表1。
表1 抑菌圈直徑測定結果(x±s,n=3,mm)
3.2 MIC和MBC測定結果
結果顯示,鞣花酸對變異鏈球菌的MIC為12.5? ? mg/mL、MBC為25 mg/mL,表明鞣花酸對變異鏈球菌有抑制作用。
3.3 細菌生物膜存活率的測定結果
結果顯示,不同質量濃度鞣花酸作用后,變異鏈球菌生物膜存活率較陰性對照組均顯著降低(P<0.01),且呈明顯劑量依賴趨勢。在MIC濃度下,變異鏈球菌生物膜的存活率僅為(16.41±1.346)%。細菌生物膜存活率測定結果見圖1。
3.4 細菌生物膜結構的觀察結果
染色結果顯示,陰性對照組中細胞互相黏附聚集形成團塊,主要呈綠色熒光,表示多為活菌,細菌生物膜結構完整。鞣花酸組中,多呈紅色熒光,表明經1/2MIC鞣花酸干預后細胞多為死菌,生物膜結構被破壞。變異鏈球菌生物膜激光共聚焦掃描圖見圖2。
3.5 細菌中EPS生成抑制率測定結果
結果顯示,鞣花酸對變異鏈球菌中水不溶性EPS和水溶性EPS的生成均有顯著抑制作用,不同質量濃度鞣花酸對2種多糖的生成抑制率均較陰性對照顯著升高(P<0.01),且該抑制作用隨著鞣花酸質量濃度的增大而增強,但鞣花酸對變異鏈球菌水不溶性EPS的抑制作用強于其對水溶性EPS的抑制作用。EPS的生成抑制率測定結果見圖3。
3.6 細菌細胞外基質中LDH活力的測定結果
結果顯示,不同質量濃度鞣花酸作用后,細菌細胞外基質中LDH活力均有所提高,且呈劑量依賴趨勢。與陰性對照組比較, 1/4MIC、1/2MIC、MIC鞣花酸組細菌細胞外基質中LDH活力均顯著升高(P<0.01)。LDH活力測定結果見圖4。
4 討論
齲病是口腔內的慢性多因素疾病,尋找安全、有效、經濟的防齲藥品一直是齲病防治研究的重點[12]。天然藥物因其毒副作用小、取材方便和經濟等特點,在齲病防治中的作用日益受到重視[13]。細菌的存在是齲病發生的第一要素,細菌在口腔中的存在形式主要為生物膜狀態,當浮游細菌黏附團聚形成生物膜后,能快速適應新的生長環境,這種由細菌和基質搭建的緊密生物膜結構,成為了阻礙藥物穿透的有力屏障[14]。在這種狀態下,生物膜結構的細菌比浮游狀態下的同種細菌耐藥性更強,并且其致毒力和對抗宿主免疫防御的能力也更強[15]。因此,在研究中除了研究浮游狀態下細菌的生長變化外,對于生物膜狀態下細菌的生長、代謝的研究顯得更加重要。EPS是細菌以蔗糖為底物分泌出的緊密圍繞菌體細胞的莢膜或者黏質層,可以參與生物膜的基質組成。其中,水不溶性EPS能夠促進細菌的蔗糖依賴性細胞黏附,加快生物膜的形成和齲病的發展;并且,其還具有生物屏障作用,可以限制生物膜內外各種物質的出入[16]。水溶性EPS是細菌在生物膜內的能量補充劑,在外源性糖缺乏時,其可以降解成單糖而提供能量[6]。LDH以高濃度存在于細菌細胞內,是變異鏈球菌中的固有酶,可以調節細菌的酸代謝[17]。由于LDH主要存在細菌細胞內,故可以通過測定細胞外基質中LDH活力來判定藥物對細菌細胞壁及細胞膜的影響[18],從而推測藥物可能的抑菌機制。因此,關于藥物對口腔內致齲細菌的作用研究需要從藥物對細菌的生長、生物膜的形成、代謝、形態結構、胞外基質的影響等多方面進行。
本研究通過打孔法抑菌圈試驗測定抑菌圈直徑,來初步判定鞣花酸對變異鏈球菌的抑制作用。結果顯示,12.5~50 mg/mL鞣花酸對變異鏈球菌產生了抑菌圈,且抑菌圈直徑>15 mm。研究中選用復方氯己定含漱液為陽性對照藥,該藥品中每500 mL中含葡萄糖酸氯己定0.6 g、甲硝唑0.1 g,為口腔常用的抑菌消炎藥[19]。本研究選用微量稀釋法測定了鞣花酸對變異鏈球菌的MIC,試驗中以刃天青顯色進行MIC的終點判定,最終得到了鞣花酸對變異鏈球菌的MIC、MBC分別為12.5、25 mg/mL,該方法比直接觀察法更加客觀、靈敏。另外,試驗結果顯示,3.125~25 mg/mL鞣花酸對變異鏈球菌的生物膜產生了抑制作用,且1/2MIC濃度的鞣花酸可破壞細菌生物膜結構。通過以上試驗結果可知,鞣花酸不僅可以抑制浮游細菌的生長,還能抑制細菌生物膜的形成。
除此之外,本研究還對鞣花酸的抑菌機制進行了研究。以1/4MIC~MIC濃度鞣花酸作用后,細菌胞外基質中LDH的活力顯著升高(P<0.01)。通過對藥物作用后細菌中EPS進行收集測定,發現鞣花酸對變異鏈球菌水不溶性EPS和水溶性EPS均有不同程度的抑制作用。通過以上研究可知,鞣花酸可以阻斷細菌毒力因子的作用途徑:第一,其可以通過抑制EPS的形成能力以降低細菌的黏附、聚集及能源供給,干擾細菌內正常的糖代謝,從而影響細菌生物膜的形成;第二,其還可以通過對變異鏈球菌的細胞產生一定程度的膜損傷,使存在于細菌胞漿內的LDH從胞漿中釋放出來,破壞細胞的正常形態和物質交換從而降低細菌活性。另外,在測定鞣花酸對EPS的抑制率時,筆者發現其對水不溶性EPS的抑制率較對水溶性EPS的高,可能是因為水不溶性EPS具有高穩定性、強黏性、不易被降解的特性,同時也是細菌具有更強致齲性的原因之一[20],因此更加說明鞣花酸能通過干擾EPS途徑抑制細菌生物膜的形成。
綜上,鞣花酸對變異鏈球菌的生長以及生物膜的形成均有抑制作用。其機制可能是通過抑制EPS產生、降低細菌的黏附、破壞細菌細胞膜,從而降低細菌活性。本研究為將鞣花酸開發成天然的防治齲病藥物提供了一定的實驗參考,但其更深入的抑菌作用及機制還有待進一步研究。
參考文獻
[ 1 ] 饒瑞瑛,任元雪,李欣瑜,等.白及提取物抗變異鏈球菌致齲效果的研究[J].中國微生態學雜志,2018,30(3):291- 295.
[ 2 ] 劉蕾,何光志,王文佳.銀杏抗菌肽蛋白基因原核表達及其重組蛋白的體內外抑菌作用研究[J].中國藥房,2019,30(18):2513-2518.
[ 3 ] VARON SE,SHRON E,ZABROVSKY A,et al. Antibacterial orthodontic cements and adhesives:a possible solution to Streptococcus mutans outgrowth adjacent to orthodontic appliances[J]. Oral Health Prev Dent,2019,17(1):251-296.
[ 4 ] 盧國英,劉建黨,
[ 5 ] 陳思佳.檸檬精油等提取物對變形鏈球菌luxS基因的影響[D].天津:天津醫科大學,2015.
[ 6 ] 羅曉東,魏麗芳,鐘昳,等.植物多酚提取物對致齲菌作用的研究進展[J].中國藥房,2019,30(1):73-77.
[ 7 ] 吳小磊,鐘晨,史金銘.鞣花酸的生物學效應[J].中國林副特產,2019,18(1):73-78.
[ 8 ] 周本宏,張紅盼,郭咸希,等.鞣花鞣質類成分研究進展[J].中國藥師,2017,20(10):1838-1841.
[ 9 ] 譚登航,王鵬嬌,張碩,等.刺梨不同藥用部位中鞣花酸的含量測定及其醇提物的體外抗氧化活性研究[J].中國藥房,2019,30(9):1236-1240.
[10] 范高福,胥振國,劉修樹,等.植物提取物鞣花酸的藥理作用及制劑研究進展[J].基因組學與應用生物學,2016,35(12):3562-3568.
[11] 徐叔云,卞如濂,陳修.藥理實驗方法學[M]. 3版.北京:人民衛生出版社,2005:1647-1662.
[12] YANG X,TOMAS BF. Tea is a significant dietary source of ellagitannins and ellagic acid[J]. J Agric Food Chem,2019,67(19):1326-1335.
[13] 宋穎,周京琳,何遠麗,等. Sortase A與變異鏈球菌致齲性關系的代謝組學研究[J].華西口腔醫學雜志,2018,36(4):360-366.
[14] XU ZT,LI MY. Comparison of inhibitions to oral bacteria among 10 Chinese herbs[J]. Chin J Mod Appl Pharm,2000,17(4):277-279.
[15] 劉曉璇,方圓,傅志華,等.植物多酚提取物對致齲菌作用的研究進展[J].中國地方病防治雜志,2019,34(2):147-149.
[16] 宋忠臣,束蓉,姚菊芳.沒食子對口腔常見細菌抑制作用的體外研究[J].口腔醫學研究,2006,22(6):634-638.
[17] SHAO D,LI J,BATES R,et al. Inhibition of gallic acid on the growth and biofilm formation of Escherichia coli and Streptococcus mutans[J]. J Food Sci,2015,80(6):299-305.
[18] KAWARAI T,NARISAWA N,YONEDA S,et al. Inhibition of Streptococcus mutans biofilm formation using extracts from Assam tea compared to green tea[J]. Arch Oral Biol,2016.DOI:10.1016/j.archoralbio.2016.04.002.
[19] 王衛,鄧倩,郭婉晴,等.洗必泰在口腔護理中應用的Meta分析[J].臨床口腔醫學雜志,2015,31(6):357-360.
[20] 吳琳,葉崢嶸,梁瑾.細菌耐藥現狀及中藥應對策略概述[J].實用中醫藥雜志,2012,28(2):159-160.
(收稿日期:2019-11-05 修回日期:2020-01-17)
(編輯:林 靜)
猜你喜歡
四川勞動保障(2021年9期)2022-01-18 05:11:08
文苑(2018年21期)2018-11-09 01:23:06
當代陜西(2018年9期)2018-08-29 01:21:00
當代陜西(2017年12期)2018-01-19 01:42:33
暨南學報(哲學社會科學版)(2016年9期)2017-01-15 13:52:00
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國衛生(2014年3期)2014-11-12 13:18:12
中國火炬(2014年4期)2014-07-24 14:22:19
