
osa-miR168a-5p靶向調節人源ADD1及E2F2基因表達的研究
2020-07-17 09:48:48陳真真蔡軍
醫學信息 2020年11期
陳真真 蔡軍
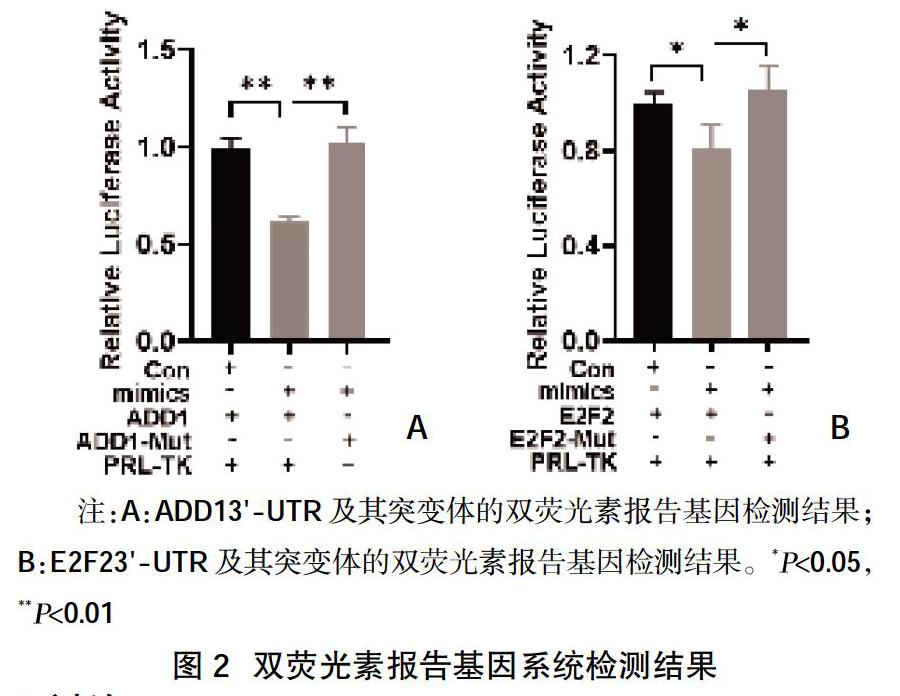
摘要:目的 ?探究植物osa-miR168a-5p靶向調節人源相關基因表達的影響。方法 ?采用TargetScan預測osa-miR168a-5p的靶基因,并篩選相關靶基因進行后續驗證。構建靶基因mRNA 3'-UTR熒光素報告基因及相應的突變體,采用雙熒光素酶報告基因活性分析系統驗證osa-miR168a-5p與靶基因的關系。結果 ?利用Targetscan預測的osa-miR168a-5p靶基因結果顯示,ADD1與E2F2mRNA3'-UTR區域均存在osa-miR168a-5p結合位點。雙熒光素酶報告基因實驗結果顯示,使用osa-miR168a-5p mimics內源性增加osa-miR168a-5p能夠抑制ADD1及E2F2報告基因的表達,將ADD1及E2F2 mRNA 3'UTR區與osa-miR-168a-5p的結合位點突變后,內源性osa-miR-168a-5p增加所引起的報告基因表達下調的作用消失。結論 ?osa-miR168a-5p能夠直接調節人源ADD1及E2F2基因表達,因此食物中的osa-miR168a進入人體很可能直接靶向人源ADD1及E2F2基因發揮相應的生物學效應。
關鍵詞:osa-miR168a;ADD1;E2F2;靶基因
中圖分類號:R3 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?文獻標識碼:A ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? DOI:10.3969/j.issn.1006-1959.2020.11.020
文章編號:1006-1959(2020)11-0064-03
Abstract:Objective ?To explore the effect of plant osa-miR168a-5p on targeted regulation of human-related gene expression.Methods ?TargetScan was used to predict osa-miR168a-5p target genes, and related target genes were screened for subsequent verification. The target gene mRNA 3'-UTR luciferin reporter gene and corresponding mutants were constructed, and the dual luciferase reporter gene activity analysis system was used to verify the relationship between osa-miR168a-5p and the target gene.Results ?The results of osa-miR168a-5p target genes predicted by Targetscan showed that both ADD1 and E2F2mRNA 3'-UTR regions had osa-miR168a-5p binding sites. The results of the dual luciferase reporter gene experiment showed that the endogenous increase of osa-miR168a-5p mimics using osa-miR168a-5p can inhibit the expression of ADD1 and E2F2 reporter genes.After mutating the binding site of ADD1 and E2F2 mRNA 3'UTR region to osa-miR-168a-5p, the effect of the endogenous increase of osa-miR-168a-5p on reporter gene expression disappeared.Conclusion ?osa-miR168a-5p can directly regulate the expression of human ADD1 and E2F2 genes. Therefore, the entry of osa-miR168a in food into the human body is likely to directly target the human ADD1 and E2F2 genes to exert the corresponding biological effects.
Key words:osa-miR168a;ADD1;E2F2;Target gene
MicroRNA(miRNA)是一類長度約為22~24個核苷酸的非編碼RNA,能夠在轉錄后層面上調控基因表達。miRNA基因在細胞核內轉錄成原始miRNA,隨后在RNAseⅢ-DROSHA的作用下形成具有發卡結構的前體miRNA,再在DICER酶的作用下切割為成熟的miRNA。成熟的miRNA的種子區域(一般為第2~8位核苷酸序列)和靶基因的3'UTR區域結合,進而抑制靶基因轉錄[1]。miRNA作為重要的轉錄后調節因子,廣泛參與心血管疾病、代謝性疾病、癌癥等疾病的發生發展[2-4]。microRNA也可以跨物種進行調節,研究發現[5],人和動物血清中始終存在外源性的植物miRNA,其中植物miR168a和miR156a表達水平最高,給予小鼠富含osa-miR168a的大米喂養后,血清及肝臟中的MIR168a水平顯著升高;進一步研究發現miR168a可以直接靶向結合低密度脂蛋白受體銜接蛋白1(LDLRAP1)外顯子,降低LDLRAP1的蛋白表達。一條microRNA可以調節多個基因的轉錄,一個基因也可以被多條microRNA調控。因此,除LDLRAP1以外,是否植物MIR168a還可以靶向其他哺乳動物的相關基因尚未可知。基于此,本研究利用生物信息學軟件預測osa-miR168a-5p的靶基因,并用雙熒光素報告基因系統分析確認,現報道如下。
1材料和方法
1.1材料與試劑 ?實驗細胞HEK-293細胞購自北京裕恒豐公司;DMEM培養基(GIBCO)、胎牛血清(GIBCO)、青霉素鏈霉素混合液(南京凱基)、PBS(Hyclone)、胰蛋白酶(GIBCO)、opti-MEM(Invitrogen)、osa-miR168amimics(廣州銳博)、Lipofectamine 2000(Invitrogen)、pmirGLO載體(優寶生物)、雙熒光素報告基因檢測試劑盒(Promega)。
1.2 HEK-293細胞培養及轉染 ?細胞在含10%胎牛血清的DMEM培養基中培養,置于5% CO2,37 ℃恒溫培養箱中孵育。轉染前1 d,胰酶消化后鋪6孔板,使其在轉染日密度為70%。轉染前首先棄掉完全培養基,加入無血清培養基,每孔細胞使用100 μl opti-MEM稀釋2 μg報告基因質粒以及100nm olosa-miR168amimics,混勻;100 μl opti-MEM稀釋2μl Lipofectamine2000轉染試劑,混勻。將上述兩種稀釋液混合后室溫放置,20 min后將其加入6孔板中,輕輕搖動混勻。轉染后的細胞在5% CO2,37 ℃恒溫培養箱中孵育6 h后更換為含10%胎牛血清的DMEM培養基。轉染48 h后,檢測報告基因活性。
1.3雙熒光素酶報告基因活性檢測 ?首先裂解細胞,配制相應的裂解液,將5X的PLB溶液用蒸餾水稀釋成1XPLB的工作液。6孔板中每孔加入500 μl 1X PLB溶液后置于水平搖床,室溫孵育15 min以便充分裂解。裂解后12000 rpm離心5 min,取上清備用。將熒光素酶檢測底物溶解于熒光素酶檢測緩沖液配置成熒光素酶檢測工作液,置于室溫。將化學發光儀設置成間隔時間2 s,測定時間10 s。取20 μl細胞裂解液上清,并加入100 μl熒光素酶檢測工作液,輕柔混勻后用化學發光儀讀數。制備停止反應液體,將試劑盒中的Stop&Glo底物溶解于Stop&Glo緩沖液,加入100 μl停止反應液后混勻并讀數。
1.4統計學方法 ?采用Graphpad Prism 8.0統計軟件進行數據分析,計量資料以(x±s)表示,行t檢驗,以P<0.05表示差異有統計學意義。
2結果
2.1預測osa-miR168a-5p靶基因及載體構建 ?利用TargetScan預測osa-miR168a-5p的靶基因,結果發現ADD1與E2F2mRNA3'-UTR區域均存在osa-miR168a-5p結合位點。osa-miR168a-5p種子區域中有連續6個核苷酸序列與人源ADD1mRNA 3'-UTR區域形成互補序列,另有連續6個核苷酸序列與人源E2F2 mRNA 3'-UTR區域形成互補序列。將ADD1,E2F2mRNA的3'-UTR與osa-miR168a-5p的種子序列的結合度進行預測,并分別以人源ADD1 mRNA和E2F2 mRNA為模板,擴增含有osa-miR168a-5p結合位點的ADD1 3'-UTR序列及含有osa-miR168a-5p結合位點的E2F2 3'-UTR序列,隨后將PCR產物通過酶切位點Nhe1和Xba1構建到pmirGLO載體上,得到pmirGLO-ADD1,pmirGLO-E2F2。對ADD1 3'-UTR及E2F2 3'-UTR 的osa-miR168a-5p結合位點進行突變后構建pmirGLO-ADD1突變質粒和pmirGLO-E2F2突變質粒,突變序列見圖1。
2.2雙熒光素報告基因系統分析確證靶基因 ?使用轉染試劑將osa-miR-168a-5pmimics與ADD13'UTRluciferase質粒或E2F23'UTR luciferase質粒以及PRL-TK熒光較準質粒共轉染HEK-293細胞后,結果顯示與對照組相比,內源性osa-miR168a-5p增加能夠降低ADD1及E2F2報告基因的表達,但當ADD1及E2F2mRNA3'UTR區osa-miR-168a-5p的結合位點突變以后,內源性osa-miR-168a-5p增加所引起的報告基因表達下調的作用消失,見圖2。
3討論
研究發現[5],哺乳動物血清及器官中存在植物miRNA,其可通過小鼠胃腸道進入機體內,提示不同種屬的miRNA可以同時在機體內共存。此外,miRNA可以跨物種進行調節,LaMonte G等[6]研究發現,瘧原蟲自身無法編碼合成miRNA,但在鐮型紅細胞中,人miR-451、miR-223及miR-19b均在瘧原蟲中富集,提示miRNA可由宿主向寄生蟲遷移。使用miRNA-451、miR223和let-7i轉染紅細胞后,顯著抑制瘧原蟲的生長,從而抵抗瘧疾的發生。Shahid S等[7]發現寄生植物菟絲子也可以通過植物miRNA影響宿主mRNA的表達。Zhou Z等[8]研究發現,在常用的抗病毒藥物金銀花煎煮后的汁液中存在大量植物microRNA2911,給予小鼠金銀花煎煮后的汁液喂養后,小鼠外周血和肺組織中的植物microRNA2911含量顯著上調,進一步研究發現microRNA2911可直接靶向甲型流感病毒H1N1,H5N1及H7N9發揮抗病毒作用。以上研究均證實miRNA可以跨越物種發揮轉錄后調節作用。miR168a是植物中常見的microRNA,番茄中的sly-miR168a能夠調節番茄的生長發育和果實成熟;擬南芥中的miR168a能夠介導對干旱、鹽堿等生物脅迫的響應[9];水稻中的miR168a(osa-miR168a)可以通過調節其靶基因AGO1的表達增加水稻種子的活力[10]。值得注意的是,miR168a是第一個被證實可以跨物種調節的植物miRNA。在哺乳動物體內,osa-miR168a可以靶向調節小鼠肝臟中的LDLRAP1,介導LDLRAP1蛋白表達沉默。目前關于osa-miR168a靶向調節其他哺乳動物基因表達的研究較少,因此對該領域的進行研究具有重要意義。
ADD1是內收蛋白基因α亞基編碼的蛋白,參與細胞膜鈉離子轉運和信號轉導[11]。研究表明[12],ADD1基因Gly460Trp位點的多態性與原發性高血壓密切相關。ADD1基因啟動子區DNA甲基化降低增加原發性高血壓的患病風險[13]。此外,ADD1還參與其他心血管疾病(動脈粥樣硬化和心肌梗死)、腎臟疾病及腫瘤的發生發展過程[11]。E2F2是E2F轉錄因子家族中的成員之一,主要參與細胞增殖的調控過程[14]。除此之外,E2F還參與細胞分化、凋亡、自噬等過程[15]。值得注意的是,E2F不僅參與腫瘤的發生發展過程,在血壓調節中也發揮重要作用,ZhouJ等[16]研究表明,E2F2全身敲除小鼠呈現高血壓;LiH等[17]研究表明,在氧化應激狀態下,靶向E2F增加二氫葉酸還原酶的含量以及恢復eNOS的功能能夠成功降低血壓。本研究使用生物信息學手段預測osa-miR168a-5p的靶基因,挑選ADD1及E2F2進行下游的驗證實驗,雙熒光素酶報告基因實驗分析顯示,使用osa-miR168a-5p mimics內源性增加osa-miR168a-5p能夠顯著抑制ADD1及E2F2報告基因的表達。然而將ADD1及E2F2 mRNA 3'UTR區與osa-miR-168a-5p的結合位點突變后,內源性osa-miR-168a-5p增加所引起的報告基因表達下調的作用完全喪失。表明人源ADD1及E2F2基因是osa-miR168a-5p的直接靶基因,證實了osa-miR168a可以調節除LDLRAP1之外的新的人源靶基因ADD1和E2F2的表達,豐富了osa-miR168a可以跨界調節的證據。
綜上所述,osa-miR168a-5p能夠直接調節人源ADD1及E2F2基因表達,食物中的osa-miR168a進入人體很可能直接靶向人源ADD1及E2F2發揮相應的生物學效應。
參考文獻:
[1]林宇鑫.MicroRNA生物標志物識別的模型構建[D].蘇州大學,2016.
[2]Thum T,Mayr M.Review focus on the role of microRNA in cardiovascular biology and disease[J].Cardiovasc Res,2012,93(4):543-544.
[3]Sethupathy P.The Promise and Challenge of Therapeutic MicroRNA Silencing in Diabetes and Metabolic Diseases[J].Curr Diab Rep,2016,16(6):52.
[4]Iorio MV,Croce CM.MicroRNA dysregulation in cancer:diagnostics,monitoring and therapeutics.A comprehensive review[J].EMBO Mol Med,2017,9(6):852.
[5]Zhang L,Hou D,Chen X,et al.Exogenous plant MIR168a specifically targets mammalian LDLRAP1:evidence of cross-kingdom regulation by microRNA[J].Cell Res,2012,22(1):107-126.
[6]LaMonte G,Philip N,Reardon J,et al.Translocation of sickle cell erythrocyte microRNAs into Plasmodium falciparum inhibits parasite translation and contributes to malaria resistance[J].Cell Host Microbe,2012,12(2):187-199.
[7]Shahid S,Kim G,Johnson NR,et al.MicroRNAs from the parasitic plant Cuscuta campestris target host messenger RNAs[J].Nature,2018,553(7686):82-85.
[8]Zhou Z,Li X,Liu J,et al.Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses[J].Cell Res,2015,25(1):39-49.
[9]Li W,Cui X,Meng Z,et al.Transcriptional regulation of Arabidopsis MIR168a and argonaute1 homeostasis in abscisic acid and abiotic stress responses[J].Plant Physiol,2012,58(3):1279-1292.
[10]Zhou Y,Zhou S,Wang L,et al.miR164c and miR168a regulate seed vigor in rice[J].J Integr Plant Biol,2020,62(4):470-486.
[11]Kiang KM,Leung GK.A Review on Adducin from Functional to Pathological Mechanisms:Future Direction in Cancer[J].Biomed Res Int,2018(2018):3465929.
[12]Zhang JR,Hu WN,Li CY.A Review of the Epidemiological Evidence for Adducin Family Gene Polymorphisms and Hypertension[J].Cardiol Res Pract,2019(2019):7135604.
[13]Zhang LN,Liu PP,Wang L,et al.Lower ADD1 gene promoter DNA methylation increases the risk of essential hypertension[J].PLoS One,2013,8(5):e63455.
[14]Infante A,Laresgoiti U,Fernandez-Rueda J,et al.E2F2 represses cell cycle regulators to maintain quiescence[J].Cell Cycle,2008,7(24):3915-3927.
[15]Muller H,Bracken AP,Vernell R,et al.E2Fs regulate the expression of genes involved in differentiation,development,proliferation,and apoptosis[J].Genes Dev,2001,15(3):267-285.
[16]Zhou J,Zhu Y,Cheng M,et al.Regulation of vascular contractility and blood pressure by the E2F2 transcription factor[J].Circulation,2009,120(13):1213-1221.
[17]Li H,Li Q,Zhang Y,et al.Novel Treatment of Hypertension by Specifically Targeting E2F for Restoration of Endothelial Dihydrofolate Reductase and eNOS Function Under Oxidative Stress[J].Hypertension,2019,73(1):179-189.
收稿日期:2020-05-12;修回日期:2020-05-22
編輯/杜帆
