羽扇豆烷型五環三萜生物活性研究進展
2020-07-23 04:23:30喬文姝趙海田
食品科學 2020年13期
喬文姝,趙海田,*,姚 磊*
(1.哈爾濱工業大學化工與化學學院,黑龍江 哈爾濱 150001;2.哈爾濱學院食品工程學院,黑龍江 哈爾濱 150086)
五環三萜類化合物是以6 個異戊二烯構成的萜類化合物,在自然界中分布廣泛[1]。因其具有多種生物功能和藥理活性,得到了食品界及醫藥領域學者的普遍關注[2-6]。按照結構不同,五環三萜可分為羽扇豆烷型、齊墩果烷型、木栓烷型和烏蘇烷型等類型。近年來,越來越多結構復雜、新穎的羽扇豆烷型三萜被發現,它們廣泛存在于樺樹皮、皺皮木瓜、酸棗仁、芍藥、甘草、毛葉棗、芒果、蘆薈、腰果等植物中[7-8],在抗腫瘤[9]、抗炎[10]、降糖[11]、護肝[12-13]、抗病毒[14]及免疫調節[15]等方面顯示出了良好的功能性,已成為天然產物五環三萜中的研究熱點。目前,常見的羽扇豆烷型五環三萜有樺木酸、樺木醇、羽扇豆醇、樺木酮酸等(圖1)。本文針對這4 種羽扇豆烷型五環三萜的抗腫瘤、抗炎、護肝、抗糖尿病生物活性及機制的最新研究進行綜述,以期為羽扇豆烷型五環三萜類化合物的深入研究和應用提供參考。
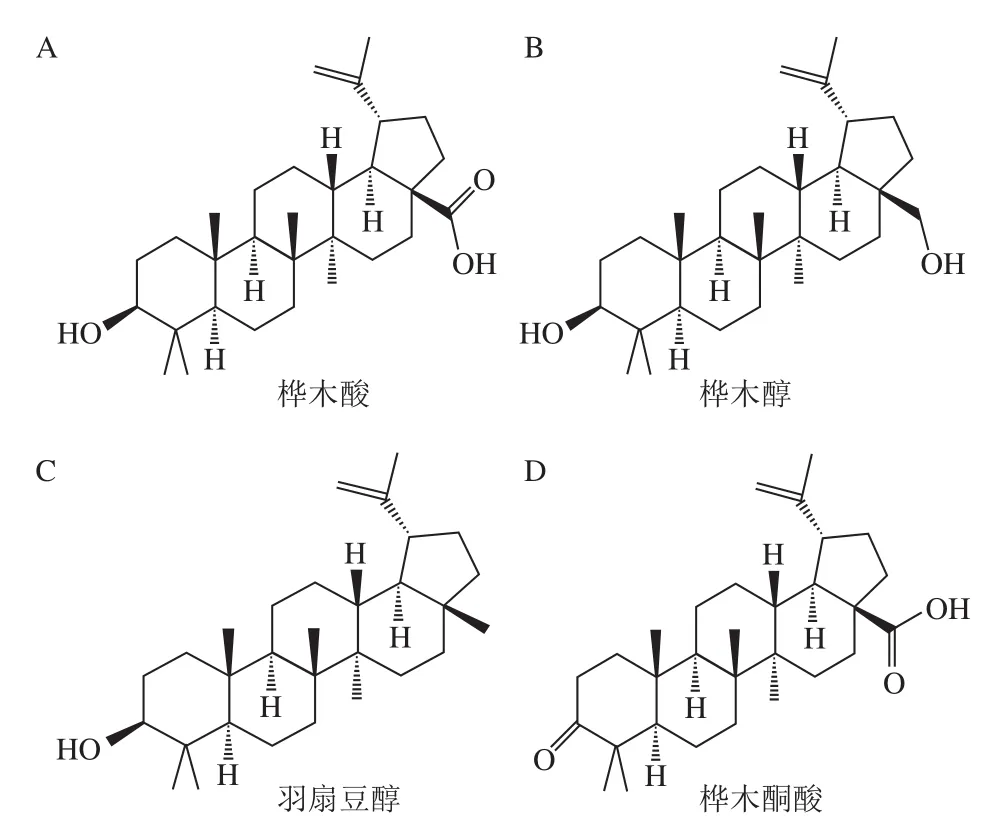
圖1 常見的羽扇豆烷型五環三萜[8]Fig. 1 Common lupinane-type pentacyclic triterpenoids[8]
1 抗腫瘤作用
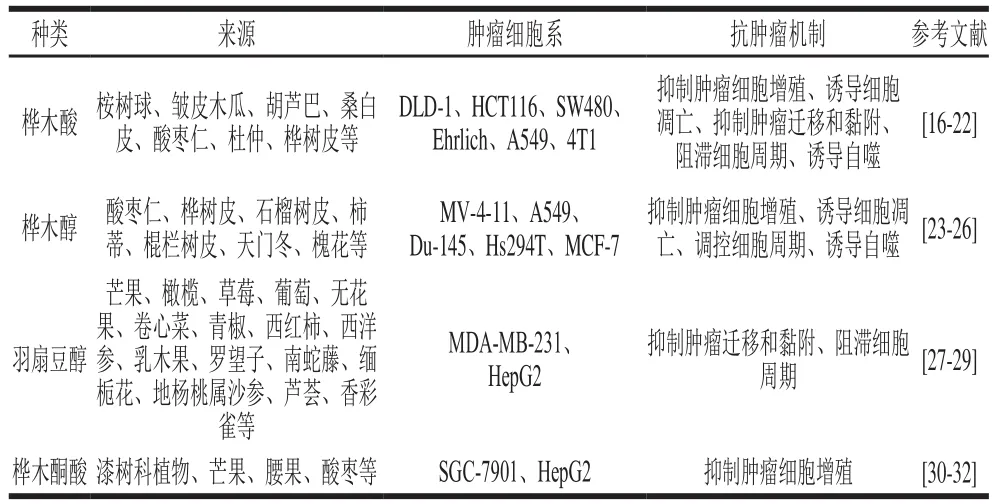
表1 羽扇豆烷型五環三萜抗腫瘤活性Table 1 Antineoplastic activity of lupinane-type pentacyclic triterpenoids
據報道,羽扇豆烷型五環三萜對結腸癌、乳腺癌、肺癌、肝癌、卵巢癌、前列腺癌、胃癌、黑色素瘤等腫瘤均有顯著的抑制作用(表1),主要的抗腫瘤作用機制有:抑制腫瘤細胞增殖、誘導腫瘤細胞凋亡、抑制腫瘤細胞遷移和黏附、調控腫瘤細胞周期、誘導腫瘤細胞自噬。
1.1 抑制腫瘤細胞增殖
抑制腫瘤細胞的增殖是羽扇豆烷型五環三萜抗癌的細胞學基礎,是其抗腫瘤作用的主要機制。Jiao Lin等[16]研究發現,乳腺癌細胞MCF-7經過樺木酸處理后,可顯著上調Caveolin-1的表達,繼而抑制磷酸化NF-κB/p65抑制蛋白(phosphorylated NF-kappa-B-inhibitor alpha,p-IκBα)表達,上調IκBα表達,增加NF-κB/p65穩定性,導致p65核轉運減少,抑制c-Myc基因的轉錄。而c-Myc表達的下調會抑制乳腺癌細胞中的有氧糖酵解,從而抑制MCF-7細胞增殖起到抗腫瘤作用(圖2)。張玲莉等[27]研究發現羽扇豆醇能通過活化Caspase通路,上調p53表達,誘導細胞凋亡及周期改變,最終抑制人高轉移肝癌HCCLM3細胞增殖,研究結果表明羽扇豆醇對肝癌的預防和治療具有一定的協同作用。Li Jizhen等[17]設計并合成了5 種樺木酸的氟化物,評估了它們對5 種腫瘤細胞系的抗增殖活性,發現它們能同時抑制拓撲異構酶I和II的活性,從而抑制癌細胞生長和增殖。Shakhtshneider等[33]利用樺木醇和阿拉伯半乳聚糖制備的復合物薄膜,對不同的癌細胞系如腹水癌細胞和肺癌細胞,均顯示出了較高的抑制作用。Wang Gang等[34]發現羽扇豆烷型五環三萜類化合物可以抑制鈣調神經磷酸酶信號通路,從而有效地抑制結腸癌細胞SW620的生長和侵襲。
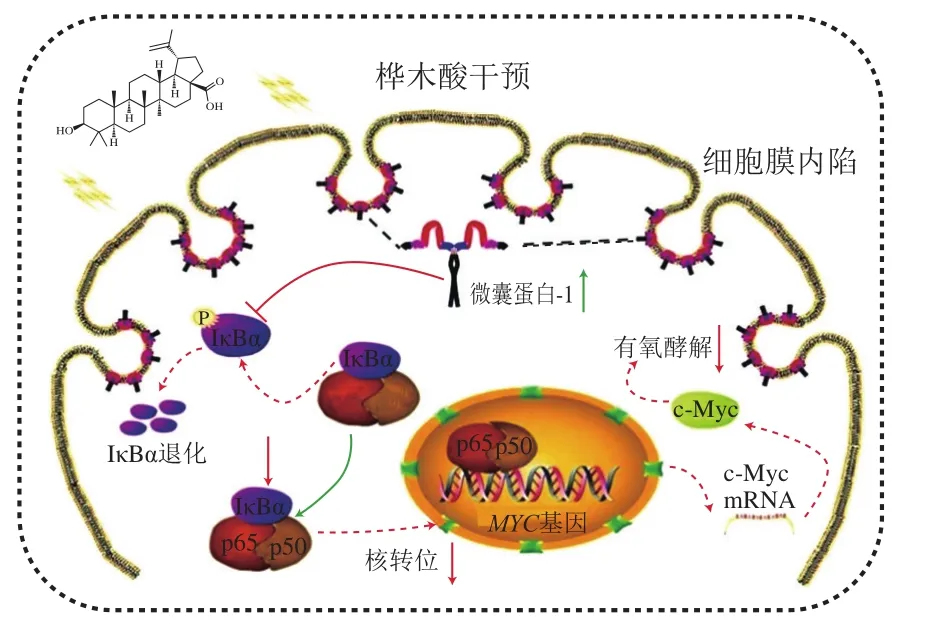
圖2 樺木酸抑制腫瘤細胞增殖作用機制[16]Fig. 2 Mechanism of action of betulinic acid in inhibiting tumor cell proliferation[16]
1.2 誘導腫瘤細胞凋亡
研究表明,羽扇豆烷型五環三萜能夠通過調節凋亡相關蛋白的表達,誘導腫瘤細胞發生凋亡,進而起到抗腫瘤作用[35]。Wang Wei等[18]發現樺木酸能上調促凋亡蛋白Bax和裂解Caspase-3的表達,下調抗凋亡蛋白Bcl-2的表達,誘導HepG2、LM3和MHCC97H等肝癌細胞凋亡并抑制癌細胞的轉移。Li Yang等[23]研究發現樺木醇可以顯著下調胃癌細胞SGC7901抗凋亡蛋白Bcl-2的表達,誘導細胞凋亡。Leong等[30]從白樺樹皮中分離提取出樺木酮酸,發現樺木酮酸可通過影響線粒體凋亡途徑,上調Bax和Bcl-2的比例,從而誘導細胞凋亡,對白血病干細胞起到抑制作用。Li Yang等[23]合成了一系列樺木醇和樺木醛的3-取代衍生物,其中,3-甲基鄰苯二甲酸酯三萜化合物顯著降低Du-145、MCF-7和Hs294T細胞中的抗凋亡蛋白Bcl-2的含量,具有對腫瘤細胞的促凋亡作用。
1.3 抑制腫瘤細胞遷移和黏附
羽扇豆烷型五環三萜可以抑制多種腫瘤細胞的遷移和黏附,有效抑制腫瘤病灶的轉移。Zeng Anqi等[19]發現樺木酸可以下調基質金屬蛋白酶(matrix metalloproteinases,MMPs)的表達并增加MMPs抑制劑(TIMP-2)的表達,顯著抑制結直腸癌細胞的遷移和侵襲,從而起到抗腫瘤作用。王明等[28]研究發現羽扇豆醇能夠下調相關蛋白COX-2、MMP-2、MMP-9的表達,從而對人乳腺癌細胞MDA-MB-231的黏附、遷移產生明顯的抑制作用。Zehra等[25]從櫟樹中分離出樺木醇,并研究了其對肺癌細胞的抗遷移活性,結果顯示,樺木醇能夠通過抑制MMP-2、MMP-9的表達,從而抑制腫瘤細胞的遷移,發揮抗腫瘤作用。Lede?i等[31]利用樺木酮酸分別與氨基、羥基、正丁基和氨基硫脲縮合,獲得了4 種官能化衍生物,發現在引入氨基后,樺木酮酸的抗腫瘤活性大大增強,能夠有效減少腫瘤病灶的轉移。
1.4 調控腫瘤細胞周期
細胞周期的阻滯是羽扇豆烷型五環三萜發揮抗腫瘤效應的重要機制[36],不同的三萜對于不同的腫瘤細胞周期的阻滯時間點不盡相同。穆曉婷等[32]研究了經樺木酮酸處理的乳腺癌細胞MCF-7和宮頸癌細胞C-33A的周期變化,結果顯示:樺木酮酸可阻滯MCF-7細胞周期在S期,阻滯C-33A細胞周期在G1-S期,從而抑制腫瘤細胞的增殖。Majeed等[20]研究發現樺木酸的衍生物可以抑制PI3K的表達,在人白血病和乳腺癌模型中將腫瘤細胞周期阻滯在G0期或G1期。Wang Ruoning等[21]制備了紫杉醇-樺木酸混合納米懸浮液,作用于乳腺癌細胞MCF-7,使其細胞周期阻滯在G0期或G1期,較少的細胞周期停滯在G2期或M期。田孜博[29]利用羽扇豆醇干預人肝癌細胞HepG2,發現羽扇豆醇可能阻斷了Wnt信號轉導通路,將HepG2的細胞周期阻滯于S期,從而抑制腫瘤細胞增殖。
1.5 誘導腫瘤細胞自噬
針對耐藥型腫瘤細胞,羽扇豆烷型五環三萜主要通過誘導細胞自噬的機制起到抗腫瘤作用[37]。Yan Xiaoning等[26]研究發現樺木醇可以上調人乳腺癌細胞MCF-7中BECLIN-1、LC 3-II的水平,同時下調P62的水平,從而誘導腫瘤細胞自噬,發揮抗腫瘤作用。Gonzalez等[22]制備了一種樺木酸的糖基化衍生物,能夠上調自噬相關蛋白ATG7、ATG5和BECLIN-1的水平,誘導腫瘤細胞自噬,起到良好的抗腫瘤效果,對樺木酸在抗腫瘤方面的應用開發具有重要意義。
羽扇豆烷型五環三萜化合物及其衍生物具有較強的、廣譜的抗腫瘤活性,在腫瘤形成的不同階段發揮著不同的作用。不僅可以引起細胞周期阻滯,抑制腫瘤細胞增殖,還可以誘導細胞凋亡或自噬,可以抑制腫瘤的形成、發展,而且也可以有效地抑制劑腫瘤細胞再生、入侵和轉移,是一種非常理想的抗腫瘤天然產物。目前,國內外學者針對樺木酸、樺木醇、羽扇豆醇等羽扇豆烷型五環三萜的抗腫瘤研究進行了大量的體內和體外實驗,并取得了良好的抗癌效果,但實際在臨床應用并具有較高價值的制劑并不多見,有待于進一步開發藥劑應用于臨床進行驗證。
2 抗炎作用
近年來,大量研究發現,羽扇豆烷型五環三萜在炎癥治療中具有良好的藥理學活性,由于其所連官能團的不同導致在藥理學和機制方面具有不同的效果,主要的抗炎作用機制有:調節細胞因子、降低氧化應激、影響花生四烯酸代謝過程。
2.1 調節細胞因子
在炎癥反應期間,免疫細胞可以分泌多種類型的細胞因子,如干擾素、白細胞介素(interleukin,IL)和腫瘤壞死因子(tumor necrosis factor,TNF)等。研究發現羽扇豆烷型五環三萜能夠調節這些細胞因子從而發揮抗炎作用。Halder等[38]制備了乳鐵蛋白修飾的負載樺木酸的聚乳酸-羥基乙酸共聚物納米顆粒,能夠上調IL-10抗炎因子,同時抑制誘導型一氧化氮合酶的表達從而產生抗炎功效。Mathew等[39]也發現樺木酸可以上調IL-10抗炎因子水平。Kasinathan等[40]研究發現羽扇豆醇可以減輕葡聚糖硫酸鈉誘導的小鼠結腸炎,其作用機制是通過降低促炎因子TNF-α、IL-1β、IL-6的水平來發揮抗炎作用。Laavola等[41]研究了樺木酸、樺木醇及其衍生物的抗炎特性,發現一種樺木醇衍生物能夠抑制IL-6細胞因子和MCP-1單核細胞趨化蛋白的表達,具有廣譜的抗炎活性。Xu Guangmen等[42]在研究樺木醇抑制脂多糖誘導的小鼠急性肝損傷時發現,樺木醇可以顯著降低血清和肝組織中IL-1β和TNF-α的水平,從而起到抗炎作用。
2.2 降低氧化應激
羽扇豆烷型五環三萜可以降低氧化應激,而氧化應激和炎癥在炎性環境下具有互相依賴的共存關系[43]。Kalra等[44]利用樺木酸緩解小鼠葡聚糖硫酸鈉誘導的結腸炎和內臟疼痛,結果顯示樺木酸可以明顯改善結腸組織中炎癥損傷等重要生化指標,如髓過氧化物酶和丙二醛的含量,同時減少血漿中亞硝酸鹽的含量,并恢復超氧化物歧化酶、過氧化氫酶和還原型谷胱甘肽水平。朱利娟等[45]發現樺木酸對地塞米松造成的氧化損傷具有防護作用,樺木酸能有效增強小鼠免疫器官中超氧化物歧化酶和還原性谷胱甘肽活性,降低丙二醛含量,從而增強免疫器官的抗氧化能力,起到抗炎作用。侯辰等[46]研究羽扇豆醇對腦缺血再灌注大鼠氧化應激損傷和炎性反應的調節作用及機制,結果表明:經羽扇豆醇干預的大鼠體內超氧化物歧化酶和還原性谷胱甘肽水平顯著升高,丙二醛水平顯著降低,炎性反應明顯減輕。
2.3 影響花生四烯酸代謝過程
在炎性環境下,花生四烯酸先通過COX-1/2形成環過氧化物,隨后在酶的催化下形成前列腺素類PGE2[47],加劇炎癥反應。羽扇豆烷型五環三萜能夠影響花生四烯酸代謝過程,減少PGE2的產生,從而起到抑制炎癥的作用。Siddiqui等[48]通過水腫和疼痛實驗,發現樺木酸可抑制人外周血單核細胞培養物中COX-2的表達,從而降低PGE2的產生量。Gon?alves等[49]研究發現新型樺木醇衍生物可以抑制IFN-γ并調節COX-2表達,影響花生四烯酸代謝過程,進而產生抗炎效應。Cao Hongmei等[50]采用脈沖超濾液相色譜-質譜法進行了環氧合酶的篩選,結果發現樺木酸能夠顯著抑制人淋巴瘤Raji細胞和人乳腺癌細胞MCF-7中COX-1的表達,起到抗炎效果。
羽扇豆烷型五環三萜可以通過調節細胞因子、減輕氧化應激、影響花生四烯酸代謝過程等機制發揮抗炎功效,并已經在炎癥治療方面顯示出令人關注的藥理特性。炎癥與動脈粥樣硬化等眾多疾病的發生相關,今后有望利用羽扇豆烷型五環三萜抗炎活性研究,進一步明確其藥理學作用機制和藥物代謝規律,設計出更合理的給藥途徑、方法和劑量,更加深入、更加廣泛地應用于臨床,使更多患者受益。
3 抗糖尿病作用
羽扇豆烷型五環三萜對糖尿病及其并發癥有一定治療作用,主要的抗糖尿病作用機制有:改善胰島功能、調節葡萄糖攝取和轉化、抑制α-葡萄糖苷酶活性。
3.1 改善胰島功能
研究表明,羽扇豆烷型五環三萜能夠通過促進胰島素分泌,增加胰島素敏感性,從而起到抗糖尿病活性。Castro等[51]研究報道,樺木酸能夠誘導ATP敏感性鉀通道(KATP)關閉,進而打開L型電壓依賴性鈣通道(L-type voltage-dependent calcium channel,L-VDCC)和鈣激活氯離子通道(calcium-activated chloride channels,CaCCs),引起鈣內流和細胞膜去極化。胞內Ca2+濃度的升高會激活蛋白激酶C(protein kinase C,PKC),PKC誘導與胰島素顆粒易位到細胞膜相關蛋白的磷酸化,從而促進胰島素顆粒移動至細胞膜并釋放胰島素。此外,樺木酸還能夠通過調控氯離子通道ClC-3,促進胰島素顆粒的酸化成熟,刺激胰島素分泌,從而起到抗糖尿病的作用(圖3)。Ahangarpour等[52]研究了樺木酸對鏈脲佐菌素-煙酰胺誘導的糖尿病小鼠中瘦素、脂聯素、肝酶水平和脂質譜的影響,發現樺木酸可以顯著降低糖尿病小鼠血漿瘦素水平、膽固醇水平和肝酶天冬氨酸氨基轉移酶水平,升高血漿脂聯素水平,從而起到改善胰島素抵抗的作用。Ramu等[53]發現在給予羽扇豆醇后,組織學觀察顯示藥物治療的糖尿病大鼠中胰島β細胞顯著再生,胰島素水平升高,血糖濃度也顯著恢復正常,這證明了羽扇豆醇能夠改善血糖和胰島β細胞再生以恢復機體正常的胰島素水平,從而起到抗糖尿病作用。Ghazanfar等[54]研究了羽扇豆醇對糖尿病大鼠代謝異常的抑制作用,發現其對大鼠的胰島β細胞具有再生和保護作用,能夠增加胰島素含量。
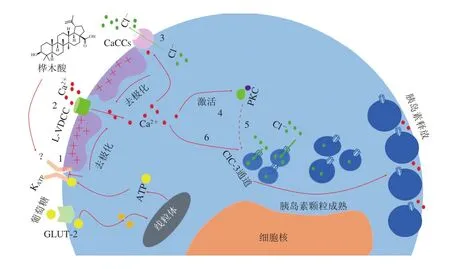
圖3 樺木酸具有促進胰島素分泌的抗糖尿病作用[51]Fig. 3 Betulinic acid plays an anti-diabetic role by promoting insulin secretion[51]
3.2 調節葡萄糖攝取和轉化
羽扇豆烷型五環三萜可以調節葡萄糖攝取和轉化(包括增加葡萄糖攝取和抑制糖異生)從而起到抗糖尿病作用。Ko等[55]發現樺木醇可以刺激3T3-L1脂肪細胞中的胰島素信號傳導,導致肝臟蛋白激酶B(protein kinase B,Akt)和絲氨酸激酶(glycogen synthase kinase 3 beta,GSK-3β)的磷酸化水平顯著升高,磷酸烯醇式丙酮酸羧激酶(phosphoenolpyruvate carboxykinase,PEPCK)的表達顯著降低,從而增強胰島素刺激的葡萄糖攝取,降低糖尿病風險。Brusotti等[56]研究發現樺木酸是PPARγ拮抗劑,能夠改善葡萄糖攝取,促進骨生成并抑制脂肪生成,可以維持胰島素敏感性,起到抗糖尿病作用。Kim等[57]研究發現樺木酸可以通過調節AMP激活的蛋白激酶(AMP activated protein kinase,AMPK)信號傳導途徑抑制肝臟糖異生,即抑制生物體將多種非糖物質轉變成葡萄糖或糖原,從而有效改善高血糖癥。
3.3 抑制α-葡萄糖苷酶活性
α-葡萄糖苷酶活性的降低可以延緩機體對碳水化合物的吸收,降低餐后血糖水平。Ding Huafang等[58]探討了樺木酸對α-葡萄糖苷酶抑制機制的新認識,采用計算機模擬分子對接技術展示了樺木酸可與α-葡萄糖苷酶活性中心的緊密結合,從而阻礙碳水化合物與酶活性中心的結合,導致α-葡萄糖苷酶活性下降,從而起到抗糖尿病的作用。Li Guoliang等[59]利用可以篩選α-葡萄糖苷酶抑制劑的熒光傳感器證實了樺木酮酸的α-葡萄糖苷酶抑制活性。Khusnutdinova等[60]則對樺木酮酸進行修飾,合成了一系列新三萜,發現這些新三萜化合物均顯示出比樺木酮酸更高的α-葡萄糖苷酶抑制活性,起到了更為顯著的抗糖尿病效果。Seong等[61]發現葛根中的羽扇豆醇可以通過抑制蛋白酪氨酸磷酸酶1B(protein tyrosine phosphatase-1B,PTP-1B)和α-葡萄糖苷酶的活性起到抗糖尿病作用。
糖尿病發病率的逐年升高,同時伴隨的慢性并發癥成為困擾人類健康的重要原因之一。羽扇豆烷型五環三萜通過改善胰島功能、調節葡萄糖攝取和轉化、抑制α-葡萄糖苷酶活性等作用機制,對I型和II型糖尿病都具有良好的治療效果,基于其抗糖尿病作用,開發出應用于糖尿病預防和治療的食品和藥物具有重要意義。但目前,羽扇豆烷型五環三萜在抗糖尿病臨床實驗數據支撐以及對羽扇豆烷型五環三萜對糖尿病并發癥的預防的研究尚鮮見報道,因此,其實際應用仍需進一步的驗證性研究。
4 護肝作用
肝臟是人體重要的代謝器官和最大的解毒器官,一旦發生病變,將對人體健康產生嚴重的影響,因此對于肝病的防護具有重要的意義。羽扇豆烷型五環三萜具有良好的護肝作用,主要的護肝作用機制有:調節相關酶活性、減緩肝臟脂質過氧化等。
4.1 調節相關酶活性
AMP激活的AMPK持續性激活,可以引發細胞的凋亡[62]。Bai Ting等[63]發現樺木醇通過影響SIRT1/AMPK信號通路,降低AMPK的表達,從而抑制大鼠肝細胞的凋亡,顯著降低乙醇誘導的小鼠酒精性肝損傷。王存琴等[64]的研究發現,羽扇豆醇可以顯著降低氧嗪酸鉀誘導的高尿酸血癥小鼠肝組織中的黃嘌呤氧化酶活性,起到保護肝臟的作用。Song Peifang等[65]發現樺木酸可以選擇性地抑制羧酸酯酶1活性,從而抑制肝臟病變,起到護肝的作用。狄英波等[66]研究發現樺木醇可以顯著抑制髓過氧化物酶、血清丙氨酸氨基轉移酶和天冬氨酸氨基轉移酶活性,從而顯著減輕肝臟病變。
4.2 減緩肝臟脂質過氧化
羽扇豆烷五環三萜能夠減緩肝臟脂質過氧化。肝臟的氧化程度過大,會引起氧化系統和抗氧化系統失衡,誘導肝組織損傷。夏偉等[67]研究樺木酸對小鼠酒精性肝損傷的保護作用,發現樺木酸能夠緩解體內乙醇代謝產物的氧化應激,降低脂質過氧化物的生成,阻止總膽固醇和總甘油三酯在肝臟的積累,減少脂肪肝的形成,從而起到護肝作用。Fernández-Martínez等[68]在薊花中提取羽扇豆醇,發現其可以減少四氯化碳誘導的急性肝損傷時的脂質過氧化從而起到保護肝臟的作用。
綜上所述,羽扇豆烷型五環三萜通過改善肝臟組織中的氧化還原體系,降低脂質過氧化物的生成,維護抗氧化系統,抑制肝損傷的發生,從而發揮有效的護肝作用。同時,羽扇豆烷型五環三萜對酒精肝、脂肪肝等肝臟疾病具有一定的緩解作用。
5 結 語
羽扇豆烷型五環三萜廣泛分布在多種植物中,種類繁多、資源豐富。近年來國內外學者研究發現一些羽扇豆烷型萜類表現出了豐富而重要的生物活性和藥理作用,除了上文提到的抗腫瘤、抗炎、抗糖尿病、護肝作用外,在抗菌、抗病毒、機體免疫調節等方面也有著巨大潛力。然而,目前羽扇豆烷型五環三萜的部分生物活性作用機制尚未明確,各種活性可能的作用靶點尚不完全清楚。此外,現有對于羽扇豆烷五環三萜的研究報道主要集中于其中的少數化合物,對其余化合物的研究還有很大的空間。
與其他天然產物抗腫瘤活性成分相比,羽扇豆烷型五環三萜最顯著的特點是其在具有廣譜抗腫瘤活性的同時,還有顯著的護肝活性。因此,考慮將羽扇豆烷型五環三萜與其他經典化療藥物聯合使用,既有可能產生協同增效作用,又可能起到降低毒副作用的特效,在未來一段時期應為相關領域研究的主要方向之一。已有研究表明,基于天然羽扇豆烷型五環三萜化合物的結構改性,有望合成出活性更強的衍生物,因此,羽扇豆烷型五環三萜改性及其構效關系研究也有很大的潛在價值。交叉學科新興技術的應用,如利用計算機模擬分子對接技術研究羽扇豆烷型五環三萜與關鍵細胞因子結合特性,對于揭示其作用機制具有很大的推進意義。相信隨著生物材料科學、藥理學、分子生物學、生物信息學等領域相關技術的不斷發展進步,羽扇豆烷型五環三萜的應用前景將更加廣闊。
