2017年中國部分地區白斑綜合征病毒3個基因的變異分析
2020-07-28 02:56:06劉曉娉劉慶慧萬曉媛
水產科學 2020年4期
劉曉娉,蔡 苗,劉慶慧,萬曉媛,黃 倢
( 1.中國水產科學研究院 黃海水產研究所,青島海洋科學與技術國家實驗室,海洋漁業科學與食物產出過程功能實驗室,農業部海水養殖病害防治重點實驗室,青島市海水養殖流行病學與生物安保重點實驗室,山東 青島 266071; 2.上海海洋大學,上海 201306 )
白斑綜合征其病原為白斑綜合征病毒(Whitespotsyndromevirus,WSSV),自20世紀90年代暴發于我國后,迅速蔓延至東南亞、南亞、南美洲、北美洲的許多國家[1-2]。白斑綜合征病毒的宿主范圍十分廣泛,苗種的攜帶是其傳播的重要來源,并且具有很高的致病性,感染白斑綜合征的病蝦死亡率高達100%[3-4]。這種廣泛的宿主范圍和高致病力被認為是白斑綜合征病毒快速和廣泛傳播的主要原因,給全球范圍內的對蝦養殖產業帶來了巨大的損失。
隨著白斑綜合征的暴發,國內外學者從病原學、傳播途徑、流行病學等方面對其展開了廣泛的研究[5]。2001年中國、美國科學家完成了其全基因組的測序工作。迄今為止,在GenBank上共有7種白斑綜合征病毒分離株的基因組全序列,分別為中國臺灣株(TW,AF440570,307 287 bp)、中國株(CN,AF332093,305 107 bp;CN01,KT995472.1,309 286 bp;CN02,KT995470.1,294 261 bp;CN03, KT995471.1,284 148 bp)、韓國株(KR,JX515788,295 884 bp)和泰國株(TH,AF369029,292 967 bp)[6]。核苷酸序列的同一性為99%,其中起源于泰國株的WSSV-TH-96-Ⅱ 具有目前最大的基因組序列,約為312 kb,被假定為祖先株[7]。白斑綜合征病毒全序列的公布,對進一步研究其基因的結構和功能起到了指導作用。白斑綜合征病毒全基因組中可變位點序列的缺失及變異可能影響其病毒的復制及傳播,產生致病力的差異。目前鑒定出5個可變基因座,包括2個具有基因組缺失的區域(ORF23/24和ORF14/15缺失區)和3個具有重復單元變化差異(VNTR)的基因座(ORF75、ORF94和ORF125)[8],主要應用于白斑綜合征病毒分子流行病學的研究。另一方面,對于白斑綜合征病毒的衣殼蛋白和囊膜蛋白功能研究也有了一定的了解[9],如VP26作為白斑綜合征病毒粒子的主要結構蛋白,將病毒粒子黏附于宿主細胞,完成進一步的侵染作用[10-11];VP28的抗血清功能可以中和白斑綜合征病毒對蝦的感染[12]。而對于病毒不同分離株其他開放閱讀框的基因變異研究相對較少。
筆者選取2017年2—6月在中國部分地區采集的44個白斑綜合征病毒陽性樣本進行白斑綜合征病毒DNA聚合酶wsv-pol、wsv313、wsv150這3個未知功能的基因位點DNA以及蛋白質序列的變異情況分析,為進一步研究白斑綜合征病毒基因變異位點的功能提供材料。
1 材料與方法
1.1 樣本來源
44份凡納濱對蝦(Litopenaeusvannamei) 白斑綜合征病毒陽性樣本于2017年2—6月分別采自河北、浙江、山東、上海、福建和廣東6個不同省市,所有樣品采集后當日冷藏運輸至實驗室于冰箱中-20 ℃冷凍保存。樣本采集信息如編號、產地、品種見表1。

表1 凡納濱對蝦樣品采集信息
(續表1)
1.2 病毒核酸提取
自保存的樣本中約取30 mg鰓組織,按照海洋動物組織基因組DNA提取試劑盒(離心柱型,天根生化科技有限公司)的說明書進行DNA提取,將提取的DNA樣本置于-20 ℃冷凍保存待用。
1.3 PCR檢測
對白斑綜合征病毒DNA樣本采用GB/T 28630.2—2012《白斑綜合征(WSD)診斷規程第2部分套式PCR檢測法》為標準進行檢測。PCR產物通過用1×TAE電泳緩沖液配制的1%的瓊脂糖凝膠進行電泳分析。
1.4 PCR擴增
用Primer 5.0軟件設計出序列的特異性引物,然后將白斑綜合征病毒陽性樣本進行wsv-pol、wsv313、wsv150基因的擴增,通過1%瓊脂糖凝膠進行電泳檢測。引物由上海生工生物工程技術服務有限公司合成,引物序列與反應條件見表2。

表2 PCR擴增的引物及反應條件
PCR擴增體系總量為25 μL,分別為:2.5 μL 10×ExTaq Buffer,2 μL dNTP,1 μL正向引物,1 μL反向引物,0.2 μL ExTaq酶,17.3 μL無菌水,1 μL DNA模板。
1.5 目的基因克隆與序列分析
將PCR產物瓊脂凝膠電泳結果中的條帶進行回收。純化的目的基因產物與載體pMD18-T連接,并轉化至TOP10感受態細胞中,接種于LB液體培養基中,37 ℃振蕩培養1 h,涂布于LB(+Amp)固體培養基上。37 ℃過夜培養,每個平板挑取至少5個單菌落接種于LB(+Amp)液體培養基中,37 ℃振蕩培養4 h。取1 μL培養物為模板按1.4中的方法進行PCR擴增,產物用1%瓊脂糖凝膠電泳檢測,檢測條帶與目的條帶一致的菌液全部送到上海生工生物工程技術服務有限公司進行DNA雙向測序。測序結果采用DNAMAN 8軟件進行序列比對分析,分別以中國大陸株(WSSV-CN,AF302293.3)序列作為參照,得出核酸水平上的變異情況;用ORF Finder處理測序結果,得到編碼的氨基酸序列,用DNAMAN 8軟件分析,得出氨基酸水平上的變異情況。
2 結果與分析
2.1 wsv-pol擴增結果
在2017年的44份白斑綜合征病毒陽性樣本中,wsv-pol的目的條帶只在福建霞美鎮、廣東湛江、山東東營和上海4個地區的樣本中出現(圖1),共有12個樣本(7、8、9、10、11、13、14、28、29、30、38、43),檢出率為27%,其余32個樣本均未擴增出該基因片段。
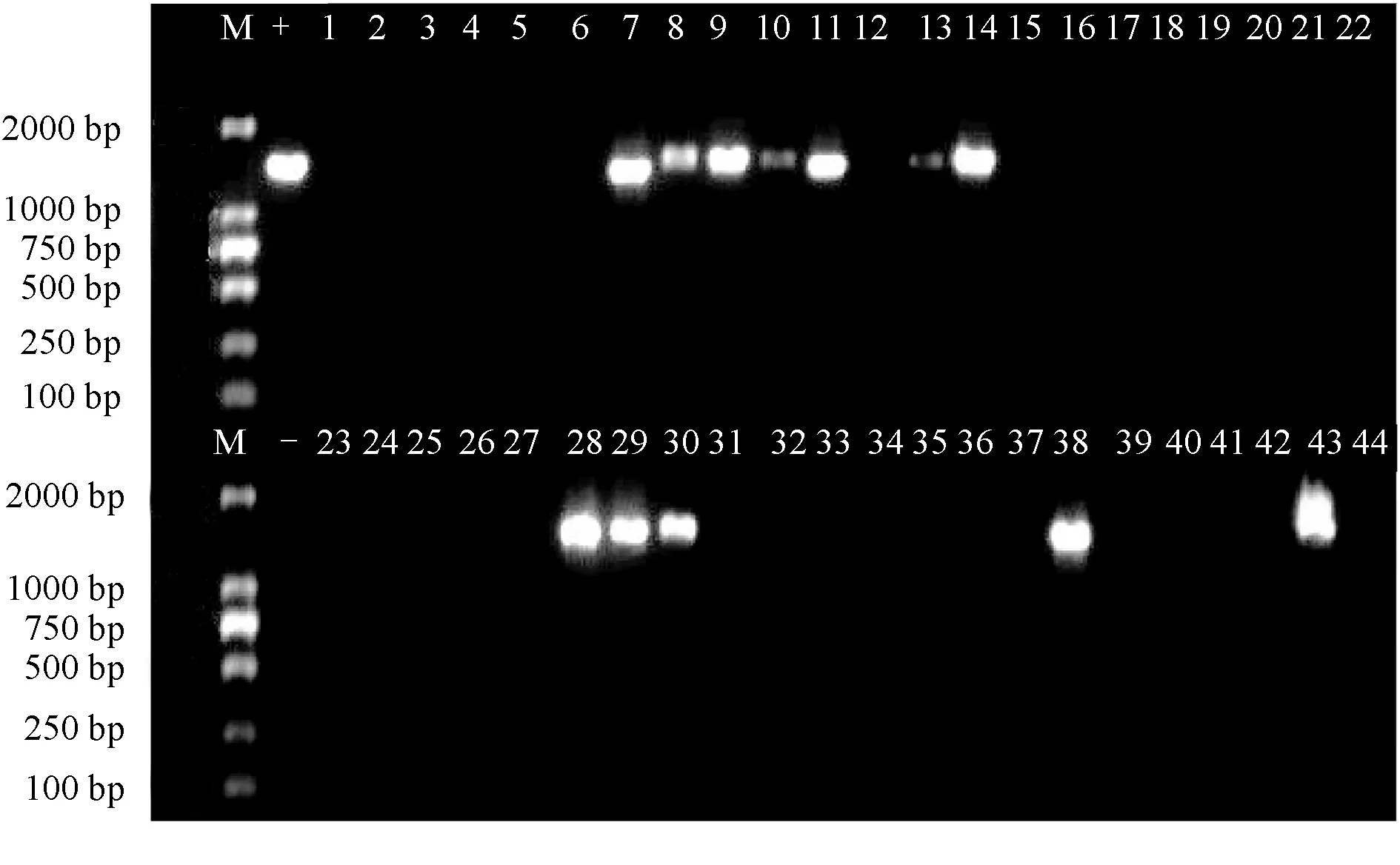
圖1 wsv-pol的PCR擴增電泳結果
2.2 wsv313擴增結果
在wsv313的擴增中,共出現12份陽性樣本,分別出現在福建深土鎮(6)、福建霞美鎮(7、8、9、10、12、13)、廣東湛江(28、29、30)、山東濱州(37)和上海(43),檢出率約為27%(圖2)。
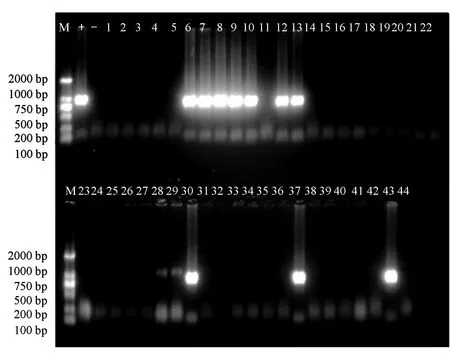
圖2 wsv313的PCR擴增電泳結果
2.3 wsv150擴增結果
在2017年44個陽性樣本的wsv150的PCR擴增中,共出現8個陽性樣本,分別出現在福建霞美鎮(7、9、11、14)、廣東湛江(28、29)、山東濱州(37)和山東東營(38) 4個地區,其檢出率為18%(圖3)。
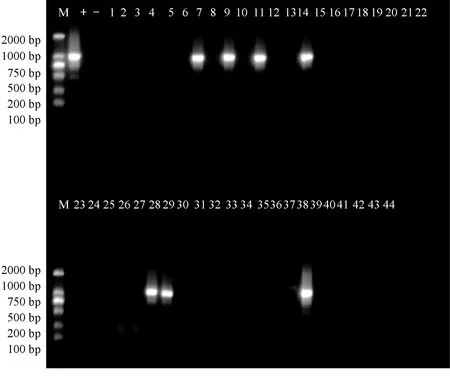
圖3 wsv150的PCR擴增電泳結果
2.4 序列比對
2017年的12個wsv-pol陽性樣本中,均在第491~493位缺失GAG 3個核苷酸,由此產生氨基酸水平上的變化是在第164位缺失1個甘氨酸(G),此外8號樣本核酸在37位T變為C,在101位 T 變為C,由此導致氨基酸在13位由酪氨酸(Y)變為組氨酸(H),在34位亮氨酸(L)變為脯氨酸(P)。10號樣本分別在106位賴氨酸(K)變為異亮氨酸(I),在214位谷氨酰胺(Q)變為精氨酸(R),在218位賴氨酸(K)變為精氨酸(R)(表3)。
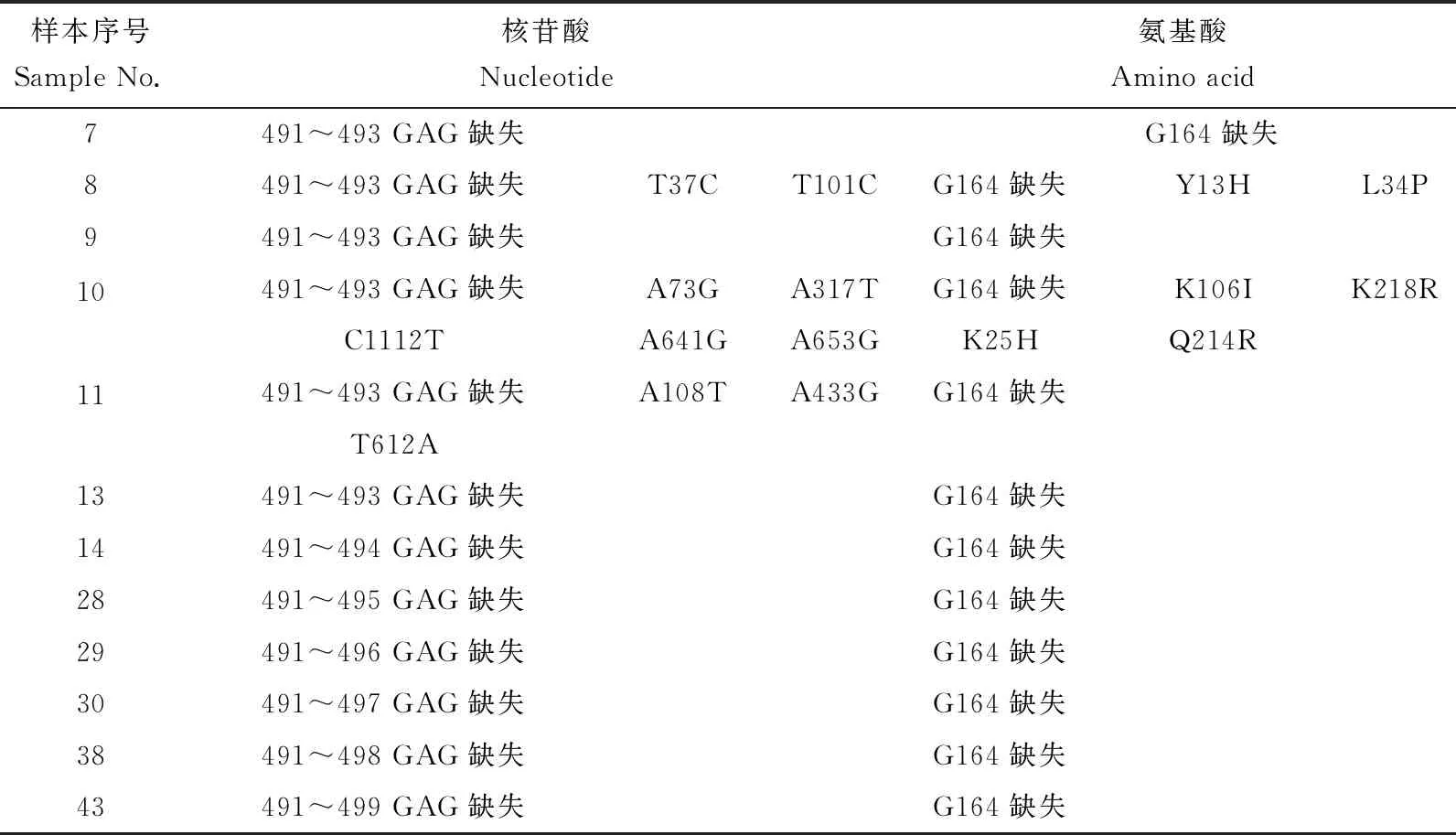
表3 wsv-pol變異情況
在2017年的wsv313的特異性擴增中共出現12個陽性樣本,均在27位核酸G替換為A。除29、30號樣本外均在822到824位插入核酸TCC,即在280位插入脯氨酸。此外9號樣本分別在86位氨基酸天冬酰胺(N)變為絲氨酸(S),在94位苯丙氨酸(F)變為絲氨酸(S),在258位精氨酸(R)變為甘氨酸(G)。而29、30號樣本,在64位天冬氨酸(D)變為甘氨酸(G)(表4)。
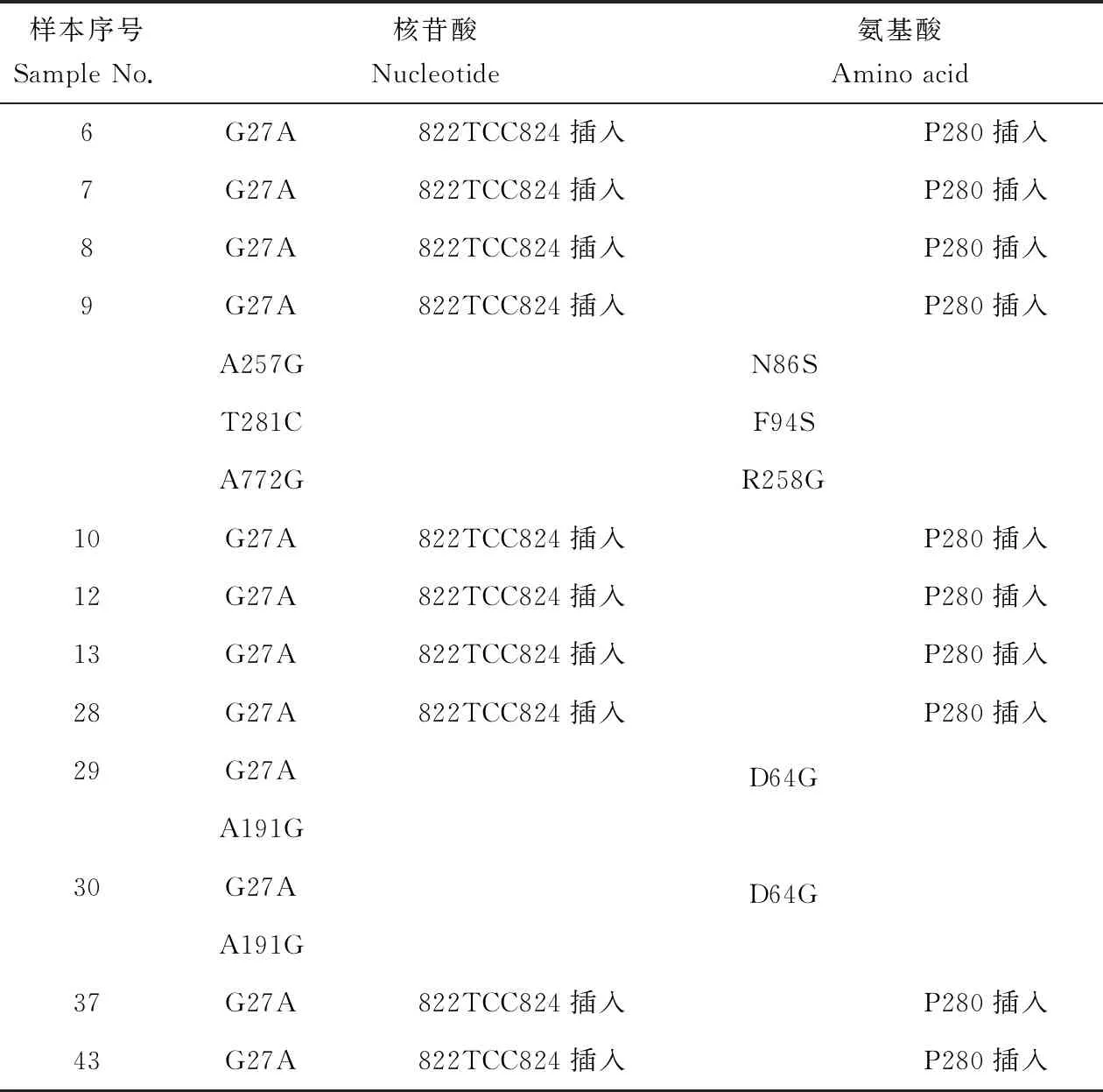
表4 wsv313變異情況
wsv150氨基酸比對結果顯示,測序樣本均出現不同程度的缺失或突變。主要存在兩大類變異情況。一是106個氨基酸大片段缺失,發生在第200個氨基酸之后(圖4),分別出現在福建霞美鎮(7、9、11、14)、廣東湛江(28、29)、山東濱州(37)、山東東營(38)的樣本中。二是發生4個氨基酸的變異也均出現在8個陽性樣本中,即第196位丙氨酸(A)突變為纈氨酸(V),第197位亮氨酸(L)突變為丙氨酸(A),第198位谷氨酰胺(Q)突變為脯氨酸(P),第199位苯丙氨酸(F)變為纈氨酸(V)(表5)。
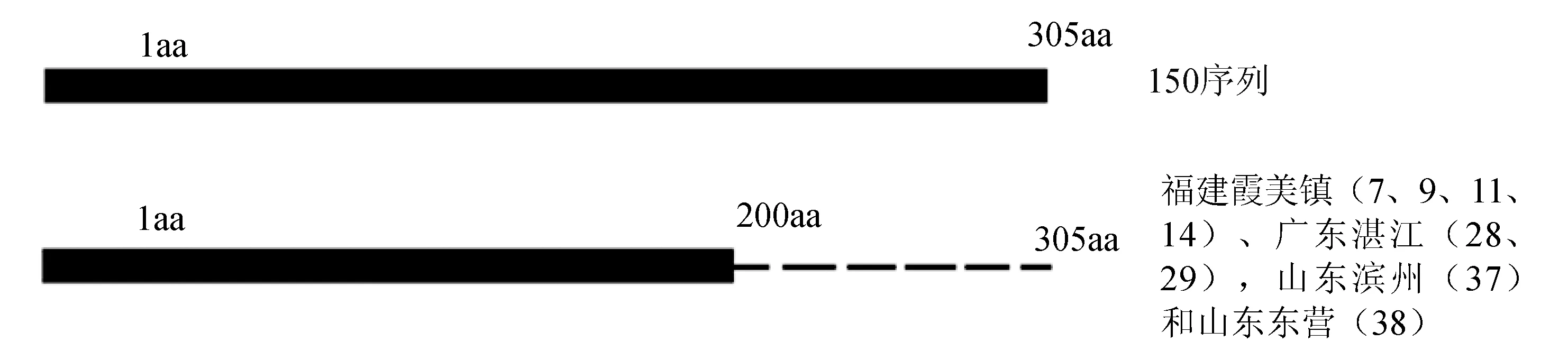
圖4 wsv150 氨基酸片段缺失示意

表5 wsv150變異情況
3 討 論
3.1 白斑綜合征病毒的基因組研究
白斑綜合征病毒是環狀雙鏈DNA病毒,基因組大小約300 kb,是目前已測序的最大動物病毒之一。白斑綜合征病毒由于地理范圍和宿主類別等環境的不同,導致不同毒株基因存在差異,這種差異往往通過研究ORF14/15和ORF23/24的缺失,ORF75、ORF94和ORF125的重復單元數目以及SNP來獲得。已有的研究表明,對蝦鰓組織是白斑綜合征病毒主要靶組織,因此試驗中取鰓組織進行病毒檢測[13]。近年的研究表明,除白斑綜合征病毒可變區外,不同分離株中個別ORF也存在明顯的變異。Li等[14]分析了不同毒力3株白斑綜合征病毒全基因組序列發現WSSV-CN01,WSSV-CN02,WSSV-CN03 與WSSV-CN相比具97.32%、97.23%、91.65%的同源性,但也存在一些差異。據報道[15],wsv001在中國不同地區的白斑綜合征病毒分離株中存在明顯變異。
3.2 白斑綜合征病毒可變位點的變異分析
DNA聚合酶是參與DNA復制的重要酶類,同時DNA聚合酶基因也是白斑綜合征病毒復制過程中關鍵作用酶基因的組成部分,屬于白斑綜合征病毒的致病因子之一[5]。完整的病毒DNA聚合酶序列全長6585 bp,編碼2195個氨基酸(wsv-pol)。本研究中,wsv-pol是一段高度保守的序列,擴增片段氨基酸位于500~950位,其擴增片段中包含DNA聚合酶的功能區。在2017年樣本的擴增中,wsv-pol的目的條帶只在福建霞美鎮、廣東湛江、山東東營和上海4個地區的樣本中出現,共有12個陽性樣本,檢出率為27%。比對結果顯示,12個陽性樣本均在第491~493位缺失GAG 3個核苷酸,由此產生的氨基酸水平上的變化是在第164位缺失1個甘氨酸。該甘氨酸的缺失對白斑綜合征病毒 DNA聚合酶功能的影響尚需進一步研究。除此之外福建霞美鎮的8、10號樣品存在不同的個別氨基酸替換現象,表現出了同一地區的差異性。研究結果顯示,wsv-pol未表現出較大程度的變異,但變異情況較為復雜,分別表現出了地區的差異性和一致性。
wsv313全長3543 bp,編碼1180個氨基酸,其195~483位氨基酸是微管結合蛋白MIP-T3的功能區。本研究側重分析wsv313編碼氨基酸功能區(195~483位氨基酸)即微管結合蛋白功能的變異情況。在福建深土鎮及霞美鎮、廣東湛江、山東東營和上海4個地區也出現12個wsv313陽性樣本,檢出率為27%。對12個樣品進行測序分析表明,wsv313均在27位核酸G替換為A,除去29、30、43號樣品外均在822到824位插入核酸TCC,即在280位插入脯氨酸。脯氨酸是一種環狀的亞氨基酸,為海洋浮游生物中一種含量居中的氨基酸,一旦進入肽鏈后,可發生羥基化作用,從而形成4-羥脯氨酸,是組成動物膠原蛋白的重要成分。有研究表明,脯氨酸可以維持膜結構穩定,清除活性氧,其累積量與植物的抗逆性呈正相關[16-17]。此外,蔡苗等[18]在對白斑綜合征病毒變異情況分析的研究中發現,wsv006位點氨基酸的主要變異情況也是脯氨酸的插入。推測白斑綜合征病毒脯氨酸的插入是為了增強自身對環境的適應性而發生的選擇性變異,上述變異是否造成微管結合蛋白功能的改變還有待進一步研究。
wsv150含305個氨基酸,比對結果顯示,測序樣品均出現不同程度的缺失或突變,主要發生在福建霞美鎮、廣東湛江、山東濱州、山東東營4個地區,檢出率約為18%。研究結果顯示,不同地區樣本變異情況存在一致性,在8個陽性樣品中均出現大片段缺失及196~199位氨基酸的突變。其大片段的氨基酸缺失可能導致蛋白質的分子構型發生變化,功能部分或者完全喪失。
綜上所述,本試驗結果表明,wsv-pol和wsv313的核酸序列保守性較高,核苷酸序列的變化引起氨基酸變化未出現較大程度的變異,各分離株間的差異也極小,而wsv150的變異程度較大,表現出地區之間高度一致性。白斑綜合征病毒不同分離株基因變異而致氨基酸變異,這些缺失和變異對致病力的影響還有待于進一步的研究。
