稻田和池塘兩種模式下金邊鯉與建鯉腸道菌群差異分析
2020-07-31 00:19:58嚴雪瑜葉香塵韋玲靜文衍紅楊著山滕忠作呂業堅
水產科學 2020年4期
關鍵詞:優勢
嚴雪瑜,葉香塵,韋玲靜,文衍紅,楊著山,滕忠作,劉 康,呂業堅
( 1.廣西壯族自治區水產引育種中心,廣西 南寧 530031; 2.柳州市漁業技術推廣站,廣西 柳州 545006; 3.柳州市融水苗族自治縣水產技術推廣站,廣西 柳州 545300 )
魚類作為一種水生動物,其自身體內及所生存的環境中都存在著大量的不同種類的微生物[1]。魚類腸道作為營養物質消化吸收的場所,其內部的微生物受到宿主腸道環境及本身遺傳特征等因素的影響,從而定殖成為腸道固有菌群[2],形成了一個腸道微生物與宿主以及所處的水生環境相互制約、相互依賴的微生態系統[3]。穩定性良好的腸道微生物環境對于魚類的消化、吸收、代謝、免疫調節和疾病防控等具有重要意義[4]。然而,魚類腸道的微生物變化是動態的,其群落組成受到宿主種類、生長發育、餌料組成和環境因素等影響[5-7]。因此,通過分析魚類腸道菌群結構上存在的差異及組成特點,可以獲得更有利于魚類生長健康的腸道微生物信息[8]。
我國稻田養魚歷史悠久,在養殖過程中,稻田里的雜草、浮萍以及浮游生物可為魚類提供充分且營養豐富的餌料,這種養殖方式在保證魚類生長和健康的同時還可改善水質以及水稻的生長條件,在一定程度上節約了養殖成本,實現資源的可持續利用[9]。因此,針對稻田養殖開發適宜的魚類養殖品種十分必要。金邊鯉(Cyprinuscarpiovar.jinbian,暫定名)因其背鰭兩側各有一條金黃色的“金邊”而得名,是由廣西水產引育種中心聯合多個單位針對稻田養殖而選育開發的鯉魚新品系[10]。金邊鯉作為一個正在培育中的新品種,其遺傳基礎、生理特征及生活習性仍需進一步研究,了解其在稻田養殖模式下的生長適應情況,查明不同養殖模式對金邊鯉腸道微生物的影響,對進一步探究金邊鯉的生長發育機制具有十分重要的意義。因此,筆者擬通過對池塘和稻田的養殖模式下金邊鯉和建鯉(C.carpiovar.jian)進行腸道宏基因組測序,分析不同鯉魚的腸道菌群差異以及不同養殖環境對鯉魚腸道菌群結構的影響,探討和評估金邊鯉在稻田養殖模式中的生長和適應情況,為金邊鯉下一步的選育工作提供研究方向和理論依據,為完善和推廣金邊鯉的稻田養殖模式提供基礎數據。
1 材料與方法
1.1 試驗設計
試驗用魚以及試驗場地均由廣西融水縣融榮水產繁殖場提供。試驗魚分別在水池中暫養7 d后隨機挑選規格整齊、體質健壯的魚各200尾,金邊鯉體質量(15.7±2.0) g,體長(8.2±0.4) cm,建鯉體質量(13.2±2.3) g,體長(7.8±0.5) cm,分別在金邊鯉和建鯉的眼瞼處注射紅色和綠色熒光標記后將2種鯉魚平均分為2組,分別同塘(同稻田)混養90 d。池塘養殖密度為45 000尾/hm2,稻田養殖密度為6000尾/hm2。試驗期間,池塘養殖日投喂人工配合飼料2次(8:30和18:00),日投喂量為魚體質量的2%;稻田養殖不投飼料。
1.2 樣品采集
試驗結束后試驗魚停食24 h,根據熒光標記分別從稻田和池塘中采集金邊鯉和建鯉各15尾,標記為池塘金邊鯉組(B1)、稻田金邊鯉組(B2)、池塘建鯉組(N1)和稻田建鯉組(N2);其中每組隨機分成3個平行組,每個平行5尾魚。按照無菌操作對樣品進行處理,用無菌生理鹽水沖洗腸管外壁,用2 mL凍存管收集腸道內容物樣品后立即放入液氮中,隨后置于冰箱-80 ℃保存備用。腸道內容物樣品用干冰保存送至上海美吉生物醫藥科技有限公司進行宏基因組文庫構建和測序。
1.3 宏基因組高通量測序及生物信息學分析
將每個平行組的腸道內容物樣品混合為一個樣品,分別對每組樣品進行MiSeq宏基因組PE文庫構建和Illumina測序。將相似度大于0.97的序列歸為1個運算分類單元(OTU),統計各樣品所含運算分類單元的數量,去除無法分類的運算分類單元后進行運算分類單元聚類分析和物種分類學分析,計算Alpha多樣性指數中的豐富度(Chao、Ace)、多樣性(Simpson、Shannon)和覆蓋率,在生物學分類的門水平和屬水平進行細菌群落結構和相對豐度分析,將標準化的操作分類單元比對京都基因和基因組百科全書(KEGG)數據庫,進行群落功能分析。
1.4 數據分析
采用Excel 2016和SPSS 22.0軟件進行數據處理,運用獨立樣本t檢驗對每個平行組的α-多樣性指數進行數據檢驗,運用單因素方差分析和Duncan′s多重比較對4組樣品的α-多樣性指數和菌群結構組成數據進行組間差異比較和顯著性分析,P<0.05表示差異顯著。
2 結果與分析
2.1 16S rRNA基因高通量測序結果分析
池塘金邊鯉組、稻田金邊鯉組、池塘建鯉組、稻田建鯉組分別產生377、250、367和455個運算分類單元,其中4組樣品共有的運算分類單元數量為175個,而每組特有的運算分類單元數量分別為39、1、15和53個。此外,金邊鯉的池塘組和稻田組產生的共有運算分類單元數量為201個,建鯉的池塘組和稻田組共有的運算分類單元數量為335個;而池塘養殖的金邊鯉和建鯉共有運算分類單元數量為282個,稻田養殖金邊鯉和建鯉的共有運算分類單元數量為240個。試驗結果表明,池塘建鯉和稻田建鯉的相同運算分類單元數量最多,而稻田金邊鯉和稻田建鯉也共享了較多相同的運算分類單元(圖1)。

圖1 各組樣品運算分類單元數量比較分析
2.2 菌群多樣性分析
在97%水平上對所有樣品進行Alpha多樣性分析,池塘金邊鯉組的Ace指數和Chao指數均高于稻田金邊鯉組,表明池塘金邊鯉的腸道菌群群落豐富度高于稻田金邊鯉;而稻田建鯉組的Ace指數和Chao指數高于池塘建鯉組,表明稻田建鯉的腸道菌群群落豐富度高于其池塘組。稻田金邊鯉組的香農指數高于池塘金邊鯉組,稻田建鯉組的香農指數亦高于池塘建鯉組,表明稻田養殖的金邊鯉及建鯉的腸道菌群群落多樣性均高于對應的池塘組;而辛普森指數結果與香農指數的結果相反,提示池塘組群落多樣性的均勻度較好。4組樣品的覆蓋率指數均達0.998以上,表明測序深度已經基本覆蓋到樣品中所有的物種(表1)。

表1 各組樣品的α-多樣性指數(n=3)
2.3 各組樣品群落組成分析
2.3.1 樣品優勢操作分類單元豐度差異分析
選取所有樣品中前50的優勢豐度操作分類單元進行差異比較分析,結果見圖2。不同養殖方式組間比較發現,建鯉的池塘組和稻田組之間的顏色相似度明顯低于金邊鯉的池塘組和稻田組的相似度,提示在稻田和池塘養殖模式下金邊鯉的腸道優勢菌群可能較建鯉的更穩定;而金邊鯉的池塘組和稻田組也存在顏色差異,表明在不同生長環境下,金邊鯉的腸道菌群結構和相對豐度各不相同。此外,同一養殖方式的組間比較發現,池塘養殖的金邊鯉和建鯉之間的顏色差異小于稻田養殖金邊鯉和建鯉之間的差異,提示池塘養殖模式下金邊鯉和建鯉的腸道優勢菌群相似性最高。試驗結果表明,不同種類及不同生長環境均會對鯉魚的腸道菌群產生一定的影響。
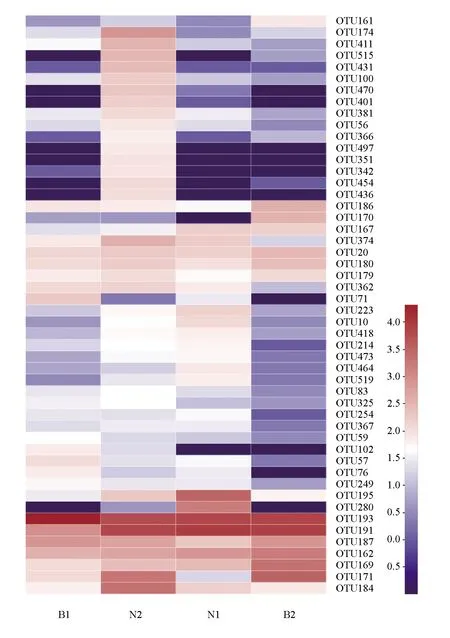
圖2 各組樣品優勢操作分類單元豐度熱圖
2.3.2 樣品菌群結構組成差異分析
在生物學分類門水平中,4組的優勢菌門主要包括變形菌門、梭桿菌門、厚壁菌門、藍細菌門、放線菌門、綠彎菌門和柔膜菌門。其中,池塘金邊鯉組中最主要的優勢菌門為變形菌門,占總數的89.56%,顯著高于其他菌門(P<0.05);其他3組的優勢菌門均為變形菌門、梭桿菌門和厚壁菌門,但細菌豐度比例存在一定的差異(圖3)。
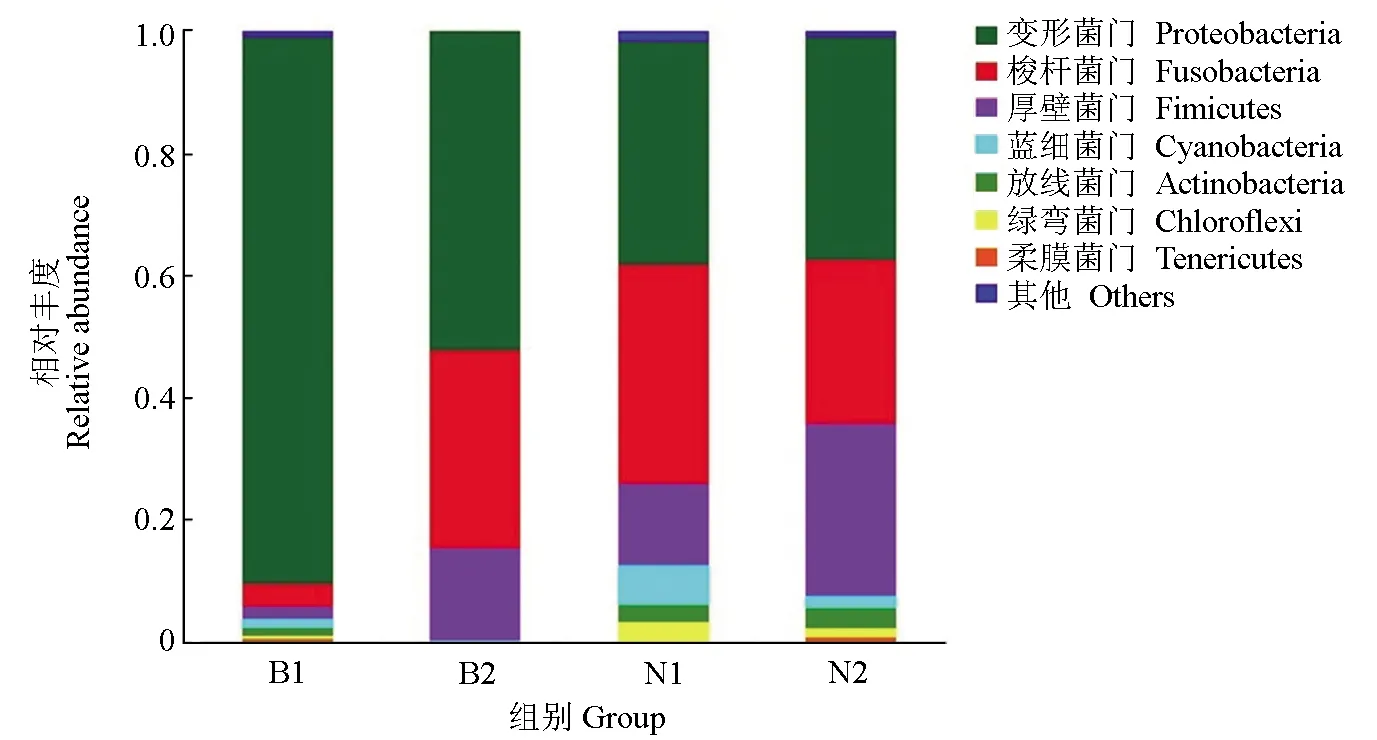
圖3 各組樣品生物分類學門水平的微生物群落結構及相對豐度
在生物學分類屬水平中,池塘金邊鯉組的第一優勢菌屬為氣單胞菌屬(82.68%),顯著高于其他菌屬(P<0.05);而稻田金邊鯉組的主要優勢菌屬包括氣單胞菌屬(Aeromonas)、鯨桿菌屬(Cetobacterium)、乳球菌屬(Lactococcus)、克雷伯菌屬(Kkbsiella)和腸桿菌屬(Enterobacter),比例分別為32.43%、32.37%、12.79%、8.36%和6.14%,5種優勢菌屬差異不顯著(P>0.05);池塘建鯉組的前3個優勢菌屬為鯨桿菌屬(35.72%)、氣單胞菌屬(27.94%)和乳球菌屬(12.29%),3種菌屬差異不顯著(P>0.05);稻田建鯉組中除去未分類和未分級的菌屬,主要優勢菌屬為鯨桿菌屬(26.83%)、氣單胞菌屬(25.16%)和乳球菌屬(8.50%),幾種優勢菌屬間差異不顯著(P>0.05)(圖4)。
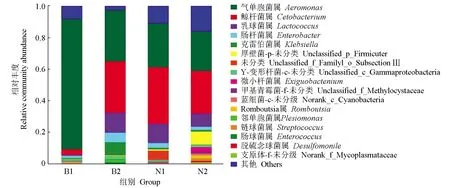
圖4 各組樣品生物分類學屬水平的微生物群落結構及相對豐度
2.4 腸道微生物16SrRNA基因功能預測分析
基于各組樣品的操作分類單元豐度水平進行腸道微生物16S rRNA基因功能預測,比對獲得了229個信號通路,分別選取每組樣品的前50個信號通路進行比較分析,其維恩圖見圖5。由圖5可見,各組樣品的腸道微生物操作分類單元共富集到45個相同的信號通路。各組樣品共有的信號通路見圖6,通過圖6可以看到其腸道微生物主要為多種物質的代謝途徑,包括丙氨酸、天冬氨酸、谷氨酸、精氨酸、脯氨酸、甘氨酸、絲氨酸、蘇氨酸、半胱氨酸、蛋氨酸、組氨酸、脂肪酸、乙醛酸、二羧酸、丙酮酸、丙酸、丁酸、煙酸、煙酰胺、甲烷、果糖、甘露糖、淀粉、蔗糖、氨基糖、核苷酸糖、甘油磷脂、卟啉、葉綠素、嘌呤、嘧啶和氮代謝,還有賴氨酸、苯丙氨酸、酪氨酸、色氨酸、纈氨酸、亮氨酸、異亮氨酸、肽聚糖、脂肪酸、葉酸、泛酸、CoA和酰基-tRNA等生物合成途徑,以及光合生物中的碳固定、原核生物中的碳固定、檸檬酸循環(TCA循環)、糖酵解/糖異生、磷酸戊糖途徑、氧化磷酸化、ABC轉運、蛋白質輸出、核糖體、磷酸轉移酶系統(PTS)和纈氨酸、亮氨酸和異亮氨酸降解等代謝通路,表明魚類腸道微生物的主要功能是參與各類物質的代謝;此外,還富集到了細菌趨化性、鞭毛組裝、細菌分泌系統、雙組分系統、同源重組和錯配修復等信號傳導、調節和修復機制途徑,提示魚類腸道微生物可能具有一定的免疫和修復調節等功能。此外,不同養殖模式組間比較發現,池塘組和稻田組建鯉的腸道微生物參與的信號通路基本一致,而池塘組和稻田組金邊鯉的腸道微生物參與的信號通路存在一定的差異,其中池塘金邊鯉組特異富集到了谷胱甘肽代謝、色氨酸代謝、β-丙氨酸代謝以及泛醌和其他萜類化合物—醌生物合成等通路,提示不同生長環境會對金邊鯉的腸道微生物的功能產生一定的影響。
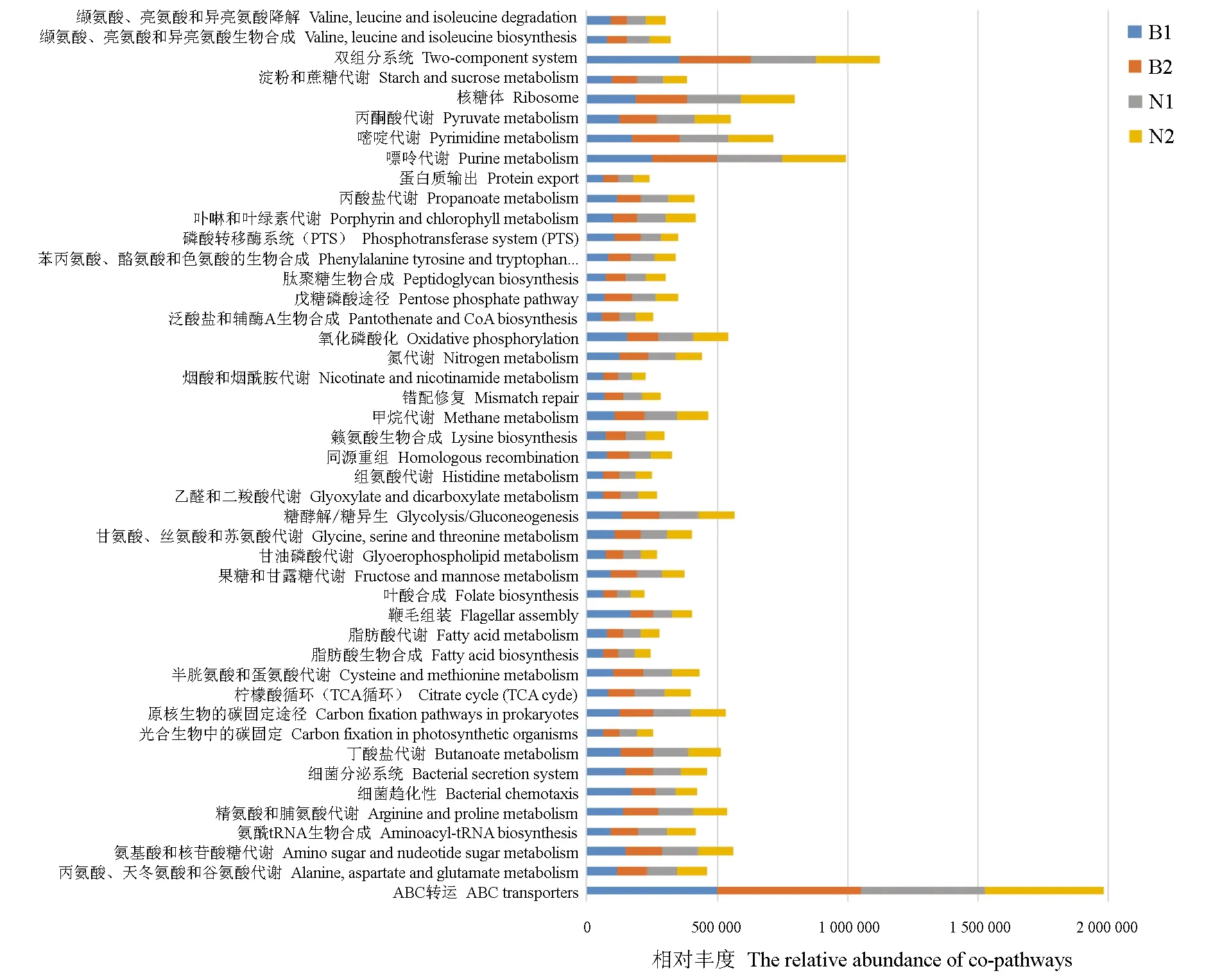
圖6 各組樣品共有信號通路比較分析
3 討 論
3.1 不同生長環境對魚類腸道菌群結構的影響
在正常的生長環境下,生物體內的大多數微生物在調節機體的各種生命功能方面起著積極的作用[11]。大量水產養殖研究表明,由于水生動物生活環境的特殊性,養殖模式、池塘水質、飼料類型等環境影響因素以及發育階段、腸道結構、營養水平和系統發育地位等變化均可打破微生物與生物之間的平衡,從而影響水生動物消化道中微生物的種類和數量[12-14]。研究證實,微生物環境可以通過水環境、飼料等引起草魚(Ctenopharyngodonidellus)腸道菌群結構發生改變[15-16]。此外,同一養殖環境下的不同魚類,其腸道微生物群落的組成還可能會受到食性、棲息地和基因型的影響[17-19]。孟曉林等[20]發現,不同質量濃度的銅可影響鯉魚腸道菌群的多樣性和結構組成,并對鯉魚腸道上皮免疫屏障造成損傷。本研究發現,不同生長環境下的金邊鯉和建鯉腸道菌群均存在一定的差異,提示養殖環境會對鯉魚的腸道微生物群落結構產生不同程度的影響。該結果與李建柱[21]的研究結果基本一致,即在魚菜共生模式下,草魚、鯽魚(Carassiusauratus)、鰱魚(Hypophthalmichthysmolitrix)和鳙魚(Aristichthysnobilis)等4種不同的鯉科魚類腸道微生物的菌落組成和優勢菌落具有一定的相似性,但是與其他養殖模式中魚類的腸道微生物菌落卻具有差異性[21]。大量研究證實,魚類的腸道菌群受多種因素的影響,但這些因素的獨立或協同作用機制有待進一步研究。
3.2 不同品種鯉魚腸道微生物優勢菌群比較分析
大量研究表明,梭桿菌門、擬桿菌門、厚壁菌門和變形桿菌門是脊椎動物腸道的核心菌群,其中變形菌門是魚類腸道微生物中最主要的優勢菌群之一[22],而擬桿菌屬(Bacteroides)、梭狀芽孢桿菌屬(Clostridium)和梭桿菌屬(Fusobacterium)中的專性厭氧菌是淡水魚的腸道優勢內源性菌群的主要種類[23]。王琴等[24]研究發現,在池塘中主養草魚混養不同比例鰱魚、鳙魚、鯽魚和匙吻鱘(Polyodonspathula)的3種混養模式下,草魚腸道菌群相似性較高,主要菌群包括變形菌門、擬桿菌門、梭桿菌門和厚壁菌門等。李建柱[21]則發現,草魚、鯽魚、鰱魚和鳙魚4種鯉科魚類腸道微生物的主要優勢菌門包括梭桿菌門、厚壁菌門、擬桿菌門、放線菌門和變形菌門,優勢菌屬包括鯨桿菌屬、梭狀芽胞桿菌屬、擬桿菌屬和芽孢桿菌屬等。王蕾[25]發現,變形菌門、藍細菌門、厚壁菌門和放線菌門等細菌門類存在于胚胎期、仔稚魚期和幼魚期的鯉魚樣本中;而混養池塘中6月齡的鯉魚腸道菌群中含量較高的菌門為變形菌門、厚壁菌門、擬桿菌門、柔膜菌門和梭桿菌門,優勢菌屬為氣單胞菌屬和乳球菌屬,鯽魚和草魚腸道菌群種類與鯉魚相似,但優勢菌和組成比例具有一定的差異。而黃進強等[26]發現,鹽堿塘養殖鯉魚腸道細菌歸屬于變形菌門、厚壁菌門、擬桿菌門和梭桿菌門,弧菌科中的氣單胞菌屬為優勢菌屬。本研究發現,金邊鯉和建鯉的主要腸道微生物均歸屬為變形菌門、梭桿菌門、厚壁菌門和藍細菌門等,主要優勢菌屬包括氣單胞菌屬、鯨桿菌屬和乳球菌屬等,且品種間無顯著差異,與其他鯉科魚類的研究結果基本一致。
3.3 金邊鯉腸道微生物優勢菌群作用分析
池塘金邊鯉組的優勢菌為變形菌門和氣單胞菌屬,占有絕對優勢;而另外3組在門分類水平和屬分類水平的菌群結構和相對豐度較為相似,優勢菌門均為變形菌門、梭桿菌門和厚壁菌門,主要優勢菌屬為鯨桿菌屬、氣單胞菌屬和乳球菌屬。其中乳球菌屬是水生動物的一類重要腸道益生菌,對宿主的免疫調節具有重要作用,可以增強宿主免疫力,其在生長過程中會產生細菌素和乳酸,抑制某些病原細菌的生長,抵抗病原菌入侵腸道[27]。而鯨桿菌屬具有發酵多肽碳水化合物的能力,并且可以產生維生素B12[28],在魚類的消化和營養過程中起到重要的作用。而在正常情況下,氣單胞菌在健康魚類的腸道中是有益的[14],該細菌可以通過阻止病原菌在腸道和養殖環境中過多繁殖,從而改善水體環境和提高宿主的免疫力[29],因此,魚類腸道中氣單胞菌為優勢菌種可能也是適應養殖環境的一種表現[26]。本研究發現,與池塘組相比,金邊鯉在稻田養殖模式下的腸道菌群結構更為豐富,并且與相同養殖條件下的建鯉無顯著差異,其優勢菌群在正常情況下基本屬于有益菌群,提示金邊鯉可以很好地適應稻田和池塘養殖環境。
4 結 論
本試驗通過比較分析稻田和池塘兩種養殖模式下的金邊鯉和建鯉腸道菌群的結構組成和豐度差異,發現稻田養殖的金邊鯉和建鯉腸道菌群多樣性高于相應的池塘養殖模式,而金邊鯉的腸道優勢菌群種類、相對豐度以及參與的信號通路與建鯉無顯著差異,提示金邊鯉與其他鯉魚一樣可適應這兩種養殖模式。
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
