龍膽苦苷對人胰腺癌細胞PANC-1凋亡及IL-6/JAK2/STAT3信號通路的影響
2020-08-16 08:31:43孟松周耀柱馬永超徐松濤金少舉
中國藥房 2020年15期
孟松 周耀柱 馬永超 徐松濤 金少舉
摘 要 目的:研究龍膽苦苷對人胰腺癌細胞PANC-1凋亡的影響,并從白細胞介素6(IL-6)/Janus激酶2(JAK2)/信號轉(zhuǎn)導與轉(zhuǎn)錄激活因子3(STAT3)信號通路角度研究其作用機制。方法:以PANC-1細胞為研究模型,采用MTT法測定0(陰性對照)、2、4、8、16、32、64、128 mg/L龍膽苦苷作用于細胞72 h后的增殖抑制率,并計算其半數(shù)抑制濃度(IC50)。分別將細胞分為陰性對照組、吉西他濱組(陽性對照,4 mg/L)和龍膽苦苷低、中、高濃度組(15、30、60 mg/L)。分別于培養(yǎng)1、3、5、7 d后,采用臺盼藍拒染法進行活細胞計數(shù),考察各組細胞的生長情況;培養(yǎng)72 h后,采用克隆形成試驗考察細胞的克隆形成率,采用Hoechst 33258染色法檢測細胞凋亡率,采用逆轉(zhuǎn)錄-聚合酶鏈式反應法和Western blotting法分別測定細胞中IL-6、JAK2、STAT3 mRNA及其蛋白表達水平。結果:4~28 mg/L龍膽苦苷均可顯著抑制細胞的增殖(P<0.05或P<0.01),并具有一定的濃度依賴性趨勢,IC50為9.54 mg/L。與陰性對照組比較,吉西他濱組和龍膽苦苷中、高濃度組活細胞計數(shù)(作用3、5、7 d)和細胞中IL-6、JAK2、STAT3 mRNA及其蛋白表達水平均顯著降低(P<0.05或P<0.01),細胞凋亡率均顯著升高(P<0.01);吉西他濱組和龍膽苦苷高濃度組細胞的克隆形成率均顯著降低(P<0.01)。與吉西他濱組比較,龍膽苦苷高濃度組細胞中上述指標水平差異均無統(tǒng)計學意義(P>0.05)。結論:30、60 mg/L龍膽苦苷可顯著抑制PANC-1細胞的增殖、誘導其凋亡,60 mg/L龍膽苦苷的作用與吉西他濱相當;其作用機制可能與抑制細胞中IL-6/JAK2/STAT3信號通路的激活有關。
關鍵詞 龍膽苦苷;人胰腺癌細胞PANC-1;凋亡;白細胞介素6;Janus激酶2;信號轉(zhuǎn)導與轉(zhuǎn)錄激活因子3;機制
ABSTRACT OBJECTIVE:To study the effects of gentiopicroside on the apoptosis of human pancreatic cancer cells PANC-1,and to explore its mechanism from the perspective of IL-6/JAK2/STAT3 signaling pathway. METHODS:Using PANC-1 cells as model,the proliferation inhibition rate of cells was tested by MTT assay after treated with 0 (negative contro),2,4,8,16,32,64,128 mg/L gentiopicroside for 72 h and IC50 were calculated. The cells were divided into negative control group,gemcitabine group (positive control, 4 mg/L) and gentiopicroside low-concentration,medium-concentration and high-concentration groups (15,30, 60 mg/L). After cultured for 1,3,5,7 d,Trypan blue exclusion staining was used to count the survival cell,and the growth of cells was investigated. After cultured for 72 h,colony formation assay was used to observe colony formation rate of cells; the apoptotic rate of cells was detected by Hoechst 33258 staining; the mRNA and protein expressions of IL-6,JAK2,STAT3 in cells were detected by RT-PCR and Western blotting assay. RESULTS:4-28 mg/L gentiopicroside could inhibit the proliferation of cells (P<0.05 or P<0.01), in concentration dependent trend; IC50 was 9.54 mg/L. Compared with negative control group, survival cell count (cultured from 3,5,7 d),mRNA and protein expressions of IL-6,JAK2 and STAT3 in cells were decreased significantly in gemcitabine group,gentiopicroside medium-concentration and high-concentration groups (P<0.05 or P<0.01), while the apoptotic rate was increased significantly (P<0.01). The colony formation rate of cells were decreased significantly in gemcitabine group and gentiopicroside high-concentration group (P<0.01). Compared with gemcitabine group,there was no statistical significance in above indexes of gentiopicroside high- concentration group (P>0.05). CONCLUSIONS:30,60 mg/L gentiopicroside could inhibit the proliferation and induce apoptosis of? PANC-1 cells,and 60 mg/L gentiopicroside is similar to gemcitabine in the effects. Its mechanism may be related to inhibiting the activation of IL-6/JAK2/STAT3 signaling pathway.
KEYWORDS Gentiopicroside; Human pancreatic cancer cells PANC-1; Apoptosis;IL-6; JAK2; STAT3; Mechanism
胰腺癌(Pancreatic cancer)是臨床上常見的消化系統(tǒng)腫瘤,具有起病隱匿、早期診斷困難、預后差及病死率高等特點,惡性程度極高[1-2]。胰腺癌發(fā)病機制復雜,目前認為炎癥反應與其發(fā)生、發(fā)展、轉(zhuǎn)移及預后密切相關,加速了疾病的進程[3-4]。白細胞介素6(IL-6)/Janus激酶2(JAK2)/信號轉(zhuǎn)導與轉(zhuǎn)錄激活因子3(STAT3)信號通路是目前發(fā)現(xiàn)的重要的炎癥信號通路之一,參與了機體的免疫調(diào)節(jié)、血管生成及細胞的增殖分化等多種生理病理過程,與腫瘤的發(fā)生、發(fā)展、轉(zhuǎn)移及侵襲等生物學行為密切相關,且其表達異常對腫瘤的預后有重要指導意義[5-6]。因此,抑制和調(diào)控IL-6/JAK2/STAT3信號通路有助于腫瘤的防治及改善預后,同時其也是篩選抗腫瘤藥物的重要靶點之一[7-8]。龍膽苦苷(Gentiopicroside)是從秦艽、龍膽草等龍膽科龍膽屬植物中提取的一種環(huán)烯醚萜苷類單體化合物,具有抗炎、抗氧化應激、抗風濕及抗肝損傷等藥理作用[9];亦有報道指出,龍膽苦苷具有抑制卵巢癌、肝癌及肺癌等腫瘤細胞的增殖、遷移等作用[10-12],但龍膽苦苷對胰腺癌的影響尚未見文獻報道。基于此,本文通過研究龍膽苦苷對胰腺癌細胞PANC-1增殖、凋亡的影響,同時檢測IL-6/JAK2/STAT3信號通路相關因子表達水平的變化,初步探討其抗胰腺癌的作用及機制,旨在為深入研究龍膽苦苷抗腫瘤藥理作用提供依據(jù)。
1 材料
1.1 儀器
CKX53型熒光倒置顯微鏡[奧林巴斯(中國)有限公司];ST-360型酶標儀(上海科華生物工程股份有限公司);Forma 系列3水套式CO2培養(yǎng)箱[賽默飛世爾科技(中國)有限公司];SJ-CJ-2FD型超凈工作臺[蘇潔醫(yī)療器械(蘇州)有限公司];GelDoc XR+型凝膠成像儀、Mini-PROTEAN? Tetra Cell型小型垂直電泳槽、小型Trans-Blot轉(zhuǎn)印槽、S1000TM 96 Deep Well 型聚合酶鏈式反應(PCR)儀[伯樂生命醫(yī)學產(chǎn)品(上海)有限公司];AE224型電子天平(上海恒平科學儀器有限公司)。
1.2 藥品與試劑
龍膽苦苷標準品[南京澤朗生物科技有限公司,批號:20180526,純度:≥98%;臨用時用含0.1%二甲基亞砜(DMSO)的培養(yǎng)基配制為相應藥液];胎牛血清(美國HyClone公司,批號:20180415);Beyozol總RNA抽提試劑、BeyoRTTM Ⅲ cDNA第一鏈合成試劑盒、Easy-LoadTM PCR Master Mix(Blue,2×)、瓊脂糖(碧云天生物技術有限公司,批號:R0011、D7178S、D7251、ST004L);Giemsa染色液(南京建成生物工程研究所,批號:180215);Hoechst 33258染色液、NP-40裂解液、二喹啉甲酸(BCA)蛋白濃度測定試劑盒、十二烷基硫酸鈉-聚丙烯酰胺(SDS-PAGE)凝膠配制試劑盒(武漢博士德生物工程有限公司,批號:AR1169、AR0107、AR0197、AR0138);兔抗IL-6、JAK2、STAT3多克隆抗體以及辣根過氧化物酶標記的羊抗兔免疫球蛋白G(H+L) 二抗(美國 Affinity Biosciences公司,批號:DF6087、AF6022、AF6294、A0001);DMEM高糖培養(yǎng)基(北京索萊寶科技有限公司);其余試劑均為分析純,水為超純水。PCR試驗所需引物由蘇州金唯智生物科技有限公司設計與合成。
1.3 細胞
人胰腺癌細胞株PANC-1購自中國科學院上海生命科學研究院細胞資源中心。
2 方法
2.1 細胞培養(yǎng)與處理
將PANC-1細胞置于含12%胎牛血清的DMEM培養(yǎng)基中,于5%CO2、飽和濕度、37 ℃條件CO2培養(yǎng)箱中培養(yǎng)(培養(yǎng)條件下同),每2~3 天更換1次培養(yǎng)液,當細胞長滿至培養(yǎng)瓶80%~95%時進行傳代。待細胞生長至對數(shù)生長期(3~4代細胞)時進行相關試驗,每項試驗均重復3次。
2.2 龍膽苦苷對PANC-1細胞增殖的影響考察
采用MTT法進行測定。收集對數(shù)生長期PANC-1細胞,以0.25%胰酶消化后制成密度為5×103個/mL的細胞懸液,按200 μL/孔的細胞密度接種于96孔板中,培養(yǎng)24 h待細胞貼壁后,分別加入終質(zhì)量濃度為0(陰性對照)、2、4、8、16、32、64、128 mg/L的含龍膽苦苷培養(yǎng)基,每個質(zhì)量濃度設3個復孔。培養(yǎng)72 h后,加入0.5% MTT溶液20 μL,繼續(xù)培養(yǎng)4 h后終止培養(yǎng)。小心吸棄培養(yǎng)孔內(nèi)上清液,然后加入DMSO 150 μL,于水平搖床上振蕩15 min使結晶物充分溶解,最后用酶標儀于490 nm波長處測定各孔吸光度(OD)值,OD值越高,則細胞活性越高。按照公式計算龍膽苦苷對PANC-1細胞的增殖抑制率:增殖抑制率(%)=(1-試驗孔OD值/陰性對照孔OD值)×100%。應用Graphpad Prism 5.0軟件計算半數(shù)抑制濃度(IC50)。
2.3 龍膽苦苷對PANC-1細胞生長的影響考察
采用臺盼藍拒染法進行測定。收集對數(shù)生長期PANC-1細胞,以0.25%胰酶消化后制成密度為1×104? ?個/mL的細胞懸液,按100 μL/孔的細胞密度接種于96孔板中。將細胞分為陰性對照組、陽性對照組和龍膽苦苷低、中、高濃度組,每組設3個復孔。待細胞貼壁后,陰性對照組細胞加入100 μL常規(guī)培養(yǎng)液,陽性對照組細胞加入終質(zhì)量濃度為4 mg/L的含吉西他濱培養(yǎng)基[13],龍膽苦苷低、中、高濃度組細胞分別加入終質(zhì)量濃度為15、30、60 mg/L的含龍膽苦苷培養(yǎng)基。于給藥培養(yǎng)第1、3、5、7 天,分別收集細胞并制成懸液,與0.4%臺盼藍溶液以9 ∶ 1(V/V)的比例混合均勻,在3 min內(nèi),用計數(shù)板計數(shù)活細胞數(shù)(顯微鏡下觀察,死細胞被染成淡藍色,而活細胞拒染)。
2.4 龍膽苦苷對PANC-1細胞克隆形成的影響考察
采用克隆形成試驗進行測定。收集對數(shù)生長期PANC-1細胞,以0.25%胰酶消化后制成細胞懸液,按500個/皿接種于60 mm培養(yǎng)皿中,按“2.3”項下方法分組、給藥,培養(yǎng)72 h。吸棄培養(yǎng)皿中培養(yǎng)液,更換為無藥新鮮培養(yǎng)基后繼續(xù)培養(yǎng),直至每個培養(yǎng)皿中均出現(xiàn)50個以上克隆即終止試驗。將培養(yǎng)皿中培養(yǎng)基小心倒棄,以磷酸鹽緩沖液(PBS)漂洗細胞3 min×2~3次,甲醇固定20 min,再用Giemsa溶液染色15 min,然后置于倒置顯微鏡下計數(shù)大于50個細胞的克隆數(shù)。按照公式計算克隆形成率:克隆形成率(%)=克隆數(shù)/接種細胞數(shù)×100%。
2.5 龍膽苦苷對PANC-1細胞凋亡的影響考察
采用Hoechst 33258熒光染色法進行測定。收集對數(shù)生長期PANC-1細胞,以0.25%胰酶消化后制成密度為2×105個/mL的細胞懸液,以2 mL/孔接種于6孔板中,按“2.3”項下方法分組、給藥,培養(yǎng)72 h。吸棄培養(yǎng)液,用無菌PBS溶液漂洗細胞3 min×2~3次,用4%多聚甲醛溶液固定30 min,然后加入Hoechst 33258染色液(5? ? ? ?μg/mL),室溫下避光染色15 min,再在超凈工作臺中風干后,于熒光倒置顯微鏡下觀察細胞凋亡情況(正常細胞核染為均勻藍色,凋亡細胞核固縮、呈強藍色致密濃染)。按照公式計算細胞凋亡率:細胞凋亡率(%)=凋亡細胞數(shù)/(凋亡細胞數(shù)+正常細胞數(shù))×100%。
2.6 龍膽苦苷對PANC-1細胞中IL-6、JAK2和STAT3 mRNA表達的影響考察
采用逆轉(zhuǎn)錄(RT)-PCR法進行測定。收集對數(shù)生長期PANC-1細胞,以0.25%胰酶消化后制成細胞懸液,按2×105個/孔接種于6孔板中,然后按“2.3”項下方法分組、給藥,培養(yǎng)72 h。吸棄培養(yǎng)液,收集細胞,用Beyozol試劑提取細胞總RNA[取2 μL總RNA樣品,于260、280 nm波長下分別檢測RNA樣品的吸光度(A),A260/A280值介于1.8~2.0之間,表明樣品純度較高]。將總RNA逆轉(zhuǎn)錄為cDNA,然后以cDNA為模板進行PCR擴增試驗。反應體系(25 μL):cDNA模板5 μL,上、下游引物各1 μL,2× Master Mix 12.5 μL,ddH2O 5.5 μL。擴增條件:94 ℃預變性3 min;94 ℃變性45 s,57 ℃退火75 s,72 ℃延伸90 s,共30個循環(huán);最后以72 ℃充分延伸10 min,結束反應。取5 μL PCR產(chǎn)物進行瓊脂糖凝膠(2%)電泳,凝膠成像儀下拍照成像,用Quantity One 4.6.2軟件分析條帶光密度值。以目的基因條帶光密度值與內(nèi)參(β-actin)條帶光密度值的比值表示目的基因mRNA的相對表達水平。待測基因引物序列及產(chǎn)物擴增長度見表1。
2.7 龍膽苦苷對細胞中IL-6、JAK2和STAT3蛋白表達的影響考察
采用Western blotting法進行測定。按“2.6”項下方法接種、分組、給藥,培養(yǎng)72 h。收集細胞于1.5 mL離心管中,加入1 mL RIPA蛋白裂解液于冰上充分裂解30 min,提取細胞總蛋白,按BCA試劑盒方法測定各樣本蛋白濃度并調(diào)整一致。然后將蛋白在95 ℃水中加熱5 min變性,80 V電壓下行SDS-PAGE電泳,300 mA恒流下將蛋白轉(zhuǎn)移至聚偏氟乙烯(PVDF)膜上;室溫下用5.0%脫脂奶粉封閉1 h,然后分別加入IL-6(1 ∶ 1 000)、JAK2(1 ∶ 1 000)、STAT3(1 ∶ 1 000)、β-actin(1 ∶ 1 000)一抗,4 ℃孵育過夜;用TBST緩沖液漂洗3 min×3次,加入二抗(1 ∶ 2 000),室溫孵育1 h;然后加入化學發(fā)光(ECL)顯色液,于凝膠成像儀下顯影拍照,用Quantity One 4.6.2軟件分析條帶灰度值。以目的蛋白條帶灰度值與內(nèi)參(β- actin)條帶灰度值的比值表示目的蛋白的相對表達水平。
2.8 統(tǒng)計學方法
應用SPSS 20.0軟件進行統(tǒng)計分析。計量資料以x±s表示,多組間樣本均數(shù)比較采用單因素方差分析,組間兩兩比較采用t檢驗。P<0.05表示差異具有統(tǒng)計學意義。
3 結果
3.1 龍膽苦苷對PANC-1細胞增殖的影響
與陰性對照比較,4、8、16、32、64、128 mg/L龍膽苦苷作用72 h后細胞的OD值均顯著降低(P<0.05或P<0.01),表明其可不同程度地抑制細胞的增殖,并且具有濃度依賴性趨勢。龍膽苦苷對PANC-1細胞的IC50為9.54 mg/L。結果見表2。
3.2 龍膽苦苷對PANC-1細胞生長的影響
在培養(yǎng)1 d后,各組的活細胞計數(shù)差異均無統(tǒng)計學意義(P>0.05)。與陰性對照組比較,在培養(yǎng)3、5、7 d后,吉西他濱組和龍膽苦苷中、高濃度組活細胞計數(shù)均明顯減少(P<0.05或P<0.01)。與吉西他濱組比較,龍膽苦苷中濃度組在培養(yǎng)3 d后的活細胞計數(shù)以及龍膽苦苷高濃度組在培養(yǎng)3、5、7 d后的活細胞計數(shù)差異均無統(tǒng)計學意義(P>0.05)。結果見表3。
3.3 龍膽苦苷對PANC-1細胞克隆形成的影響
與陰性對照組比較,吉西他濱組和龍膽苦苷高濃度組細胞的克隆形成數(shù)顯著減少(P<0.01),克隆形成率顯著降低(P<0.01)。與吉西他濱組比較,龍膽苦苷高濃度組細胞的克隆形成數(shù)和克隆形成率差異均無統(tǒng)計學意義(P>0.05)。結果見圖1、表4。
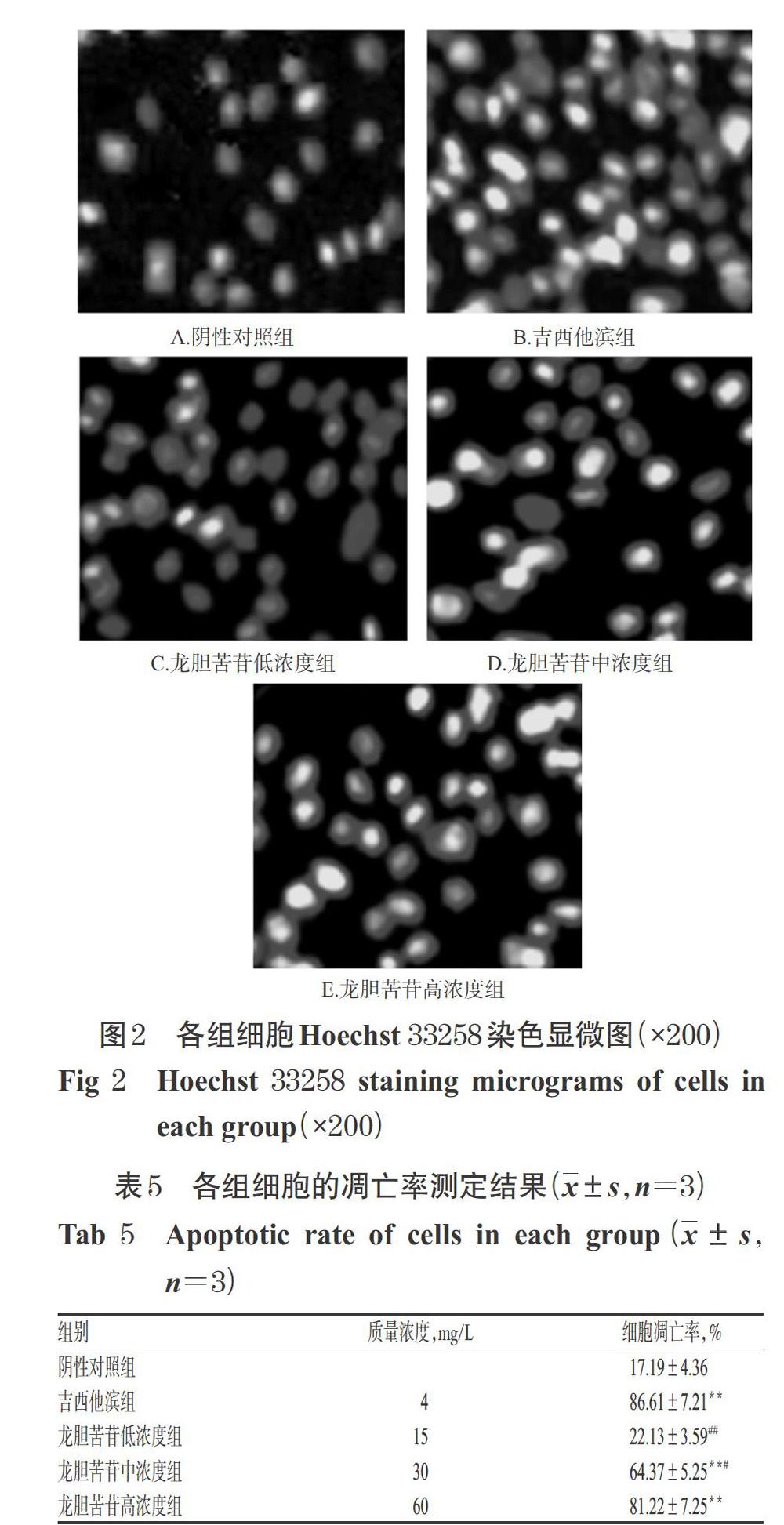
3.4 龍膽苦苷對PANC-1細胞凋亡的影響
與陰性對照組比較,吉西他濱組和龍膽苦苷中、高濃度組細胞凋亡率顯著升高(P<0.01)。與吉西他濱組比較,龍膽苦苷高濃度組細胞凋亡率差異無統(tǒng)計學意義(P>0.05)。結果見圖2、表5。
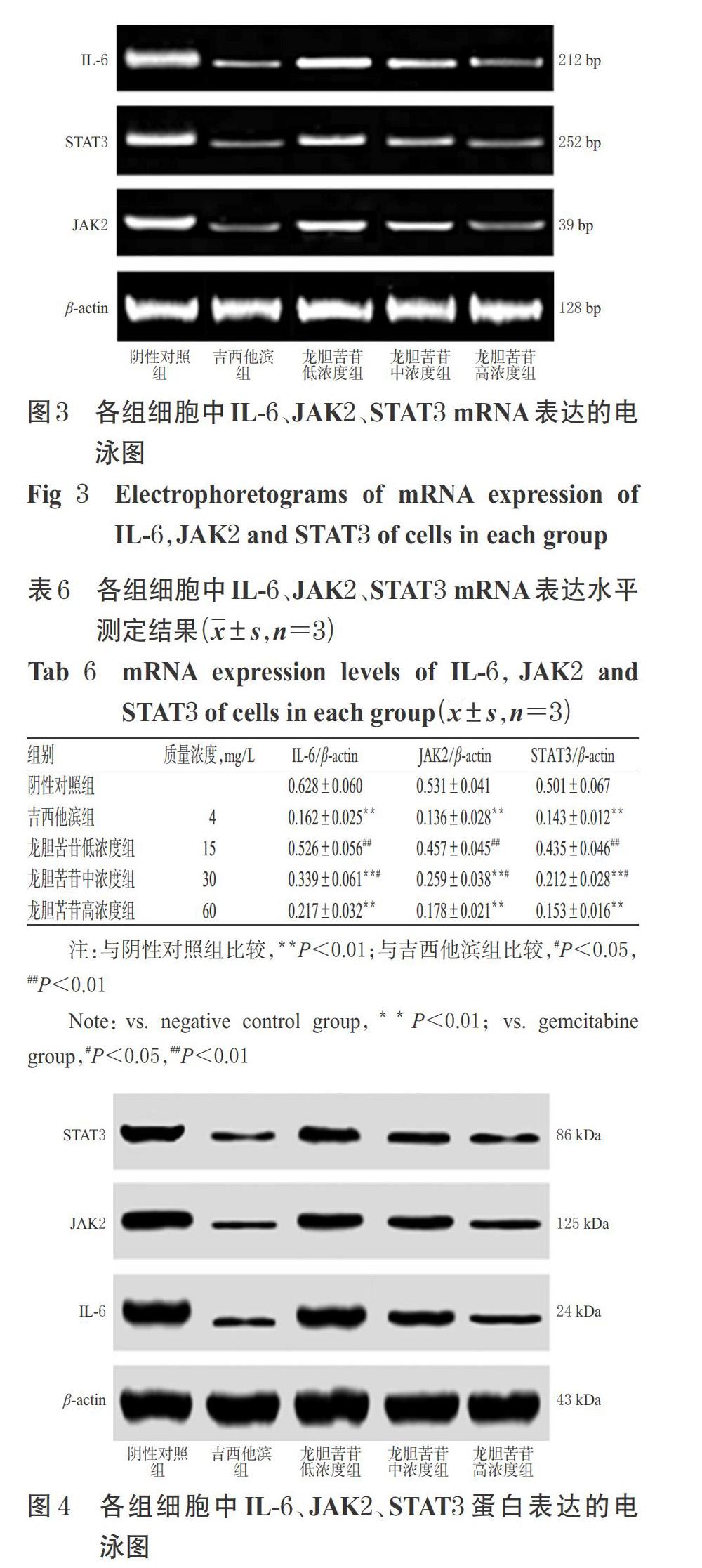
3.5 龍膽苦苷對PANC-1細胞中IL-6、JAK2、STAT3 mRNA表達水平的影響
與陰性對照組比較,吉西他濱組和龍膽苦苷中、高濃度組細胞中IL-6、JAK2、STAT3 mRNA表達水平顯著降低(P<0.01)。與吉西他濱組比較,龍膽苦苷高濃度組細胞中IL-6、JAK2、STAT3 mRNA表達水平差異均無統(tǒng)計學意義(P>0.05)。結果見圖3、表6。
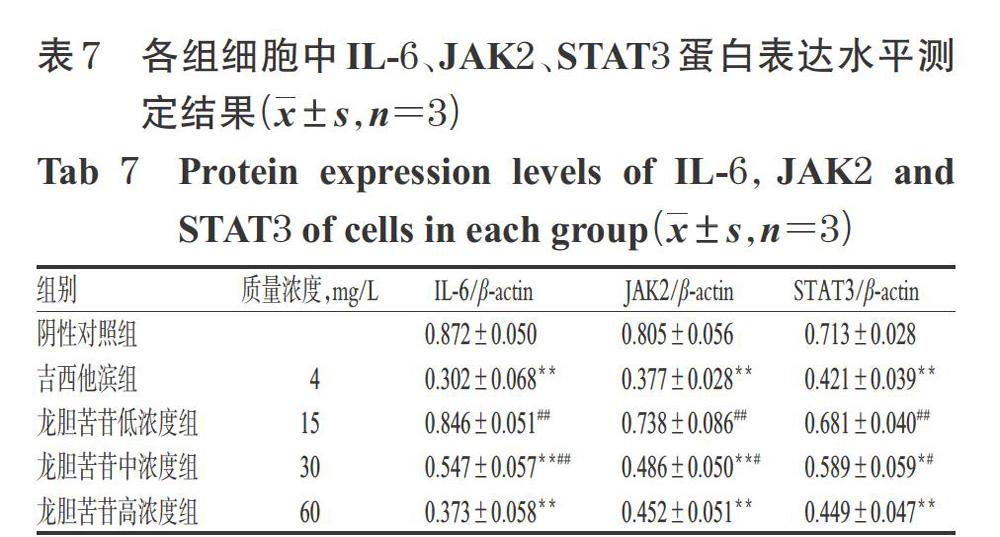
3.6 龍膽苦苷對PANC-1細胞中IL-6、JAK2、STAT3蛋白表達水平的影響
與陰性對照組比較,吉西他濱組和龍膽苦苷中、高濃度組細胞中IL-6、JAK2、STAT3蛋白表達水平顯著降低(P<0.05或P<0.01)。與吉西他濱組比較,龍膽苦苷高濃度組細胞中IL-6、JAK2、STAT3蛋白表達水平差異均無統(tǒng)計學意義(P>0.05)。結果見圖4、表7。
4 討論
炎癥信號通路IL-6/JAK2/STAT3在腫瘤的發(fā)生、發(fā)展過程中發(fā)揮著至關重要的作用[14]。臨床研究發(fā)現(xiàn),胰腺癌患者腫瘤組織中IL-6、JAK2、STAT3等的表達異常升高,且與腫瘤的惡性程度及預后不良呈正相關[15]。其中,IL-6是目前研究較多且功能廣泛的細胞因子之一,其參與了炎癥反應、免疫調(diào)控、細胞分化及增殖等生理病理過程,其表達升高可促進腫瘤的發(fā)生、發(fā)展、遷移及侵襲等生物學行為,也是判斷惡性腫瘤預后的重要指標之一[16]。IL-6與其靶細胞上相應的IL-6受體結合后,激活細胞膜表面的糖蛋白130(GP130),從而激活與GP130關聯(lián)的JAK,促進受體蛋白酪氨酸激酶活化,并與STAT3結合,使STAT3發(fā)生磷酸化激活后,誘導核因子? κB(NF-κB)從NF-κB/NF-κB抑制蛋白(IκB)復合體上解離為游離的磷酸化狀態(tài),然后通過核膜進入細胞核內(nèi),與其相應的DNA反應元件結合,調(diào)控炎癥、凋亡或增殖相關因子的表達,促進細胞增殖、抑制細胞凋亡,參與到腫瘤細胞的增殖、遷移及侵襲等生物學行為中[17-20]。因此,采取有效治療措施下調(diào)IL-6/JAK2/STAT3信號通路的表達有助于腫瘤的防治。本研究發(fā)現(xiàn),30、60 mg/L龍膽苦苷可顯著抑制PANC-1細胞的增殖、生長和克隆形成,促進細胞凋亡,下調(diào)細胞中IL-6、JAK2、STAT3 mRNA及其蛋白表達,并且60 mg/L龍膽苦苷的作用與4 mg/L吉西他濱的作用相近(吉西他濱是臨床上治療胰腺癌的基礎化療藥物[21],故在本研究中將其設為陽性對照)。
綜上所述,龍膽苦苷對人胰腺癌細胞PANC-1具有抑制增殖、誘導凋亡的作用,其機制可能與抑制IL-6/ JAK2/STAT3信號通路的激活有關,但其更多的作用機制后續(xù)需進一步研究。
參考文獻
[ 1 ] ABBASSI R,SCHMID RM. Evolving treatment paradi- gms for pancreatic cancer[J]. Visc Med,2019,35(6):362-372.
[ 2 ] 劉夢奇,吉順榮,徐曉武,等. 2019年胰腺癌研究及診療新進展[J].中國癌癥雜志,2020,30(1):1-10.
[ 3 ] WANG Y,YANG G,YOU L,et al. Role of the micro- biome in occurrence,development and treatment of pancreatic cancer[J]. Mol Cancer,2019,18(1):173-186.
[ 4 ] STONE ML,BEATTY GL. Cellular determinants and therapeutic implications of inflammation in pancreatic cancer[J]. Pharmacol Ther,2019,201(9):202-213.
[ 5 ] ZHANG X,LU H,HONG W,et al. Tyrphostin B42 atte- nuates trichostatin A-mediated resistance in pancreatic cancer cells by antagonizing IL-6/JAK2/STAT3 signaling[J]. Oncol Rep,2018,39(4):1892-1900.
[ 6 ] LIU X,WANG J,WANG H,et al. REG3A accelerates pancreatic cancer cell growth under IL-6-associated inflammatory condition:involvement of a REG3A-JAK2/STAT3 positive feedback loop[J]. Cancer Lett,2015,362(1):45-60.
[ 7 ] ZHAO H,GUO Y,LI S,et al. A novel anti-cancer agent icaritin suppresses hepatocellular carcinoma initiation and malignant growth through the IL-6/JAK2/STAT3 pathway[J]. Oncotarget,2015,6(31):31927-31943.
[ 8 ] 劉建民,黃良文,朱旭紅,等. 3種活血化瘀中藥復方含藥血清對神經(jīng)膠質(zhì)瘤U251細胞JAK/STAT信號通路的影響[J].中國藥房,2017,28(16):2176-2179.
[ 9 ] 謝小倩,李賀,王亞樂,等.龍膽苦苷對佐劑性關節(jié)炎大鼠的抗炎作用[J].中國實驗方劑學雜志,2020,26(6):58- 63.
[10] XIE X,LI H,WANG Y,et al. Therapeutic effects of gen- tiopicroside on adjuvant-induced arthritis by inhibiting? ?inflammation and oxidative stress in rats[J]. Int Immu-? ?nopharmacol,2019. DOI:10.1016/j.intimp.2019.105840.
[11] 王美靈,張雷明,郝妍斐,等.龍膽苦苷對類風濕性關節(jié)炎的保護作用及其機制[J].中國藥理學與毒理學雜志,2019,33(9):721.
[12] LI X,YANG C,SHEN H. Gentiopicroside exerts convincing antitumor effects in human ovarian carcinoma cells (SKOV3) by inducing cell cycle arrest,mitochondrial? mediated apoptosis and inhibition of cell migration[J]. J Buon,2019,24(1):280-284.
[13] 梁春麗,王崢,李炳,等.姜黃素對人胰腺癌PANC-1細胞甲基化轉(zhuǎn)移酶表達影響的體外研究[J].上海中醫(yī)藥雜志,2015,49(10):77-79、97.
[14] KIM JH,CHOI HS,KIM SL,et al. The PAK1-Stat3 signaling pathway activates IL-6 gene transcription and human breast cancer stem cell formation[J]. Cancers:Basel,2019,11(10):1527-1546.
[15] RAZIDLO GL,BURTON KM,MCNIVEN MA.Interleukin-6 promotes pancreatic cancer cell migration by rapidly activating the small GTPase CDC42[J]. J Biol Chem,2018,293(28):11143-11153.
[16] 陳濤,曾永鴻.腫瘤微環(huán)境中異常表達的白介素6對腫瘤進程的影響[J].解剖學雜志,2016,39(4):501-503.
[17] 任為,牟宜雙,許可,等.迷迭香酸通過誘導凋亡及抑制NF-κB信號通路發(fā)揮抗肝癌作用的機制研究[J].中藥藥理與臨床,2016,32(2):31-35.
[18] 張曼澤,李峰生,王思念,等. IL-6通過JAK2/STAT3信號通路促進CD133-肺腺癌細胞(A549)獲得干性特征[J].軍事醫(yī)學,2018,42(9):678-683.
[19] LIU Z,CHEN T,LU X,et al. Overexpression of variant PNPLA3 gene at I148M position causes malignant transformation of hepatocytes via IL-6-JAK2/STAT3 pathway in low dose free fatty acid exposure:a laboratory investigation in vitro and in vivo[J]. Am J Transl Res,2016,8(3):1319-1338.
[20] 李雷雷,郭彬,郭佳培,等.腫瘤相關巨噬細胞通過JAK2/STAT3途徑調(diào)控肝癌細胞凋亡的機制研究[J].現(xiàn)代預防醫(yī)學,2017,44(13):2406-2410.
[21] 彭夢媛,邱峰,黃丹,等.姜黃素對胰腺癌SW1990細胞耐吉西他濱的逆轉(zhuǎn)作用及機制研究[J].中國藥房,2019,30(9):1192-1197.
(收稿日期:2020-02-24 修回日期:2020-06-12)
(編輯:林 靜)
