毛竹擴張對亞熱帶常綠闊葉林主要樹種葉結構型性狀的影響
2020-08-18 07:24:56鐘雅琪鐘全林李寶銀徐朝斌程棟梁樂新貴鄭文婷
生態學報 2020年14期
鐘雅琪,鐘全林,,*,李寶銀,余 華,4,徐朝斌,程棟梁,,樂新貴,鄭文婷,6
1 福建師范大學地理科學學院, 福州 350007
2 濕潤亞熱帶山地生態國家重點實驗室培育基地, 福州 350007
3 福建師范大學福建省植物生理生態重點實驗室, 福州 350007
4 閩江學院海洋學院, 福州 350108
5 江西陽際峰國家自然保護區管理局, 貴溪 335400
6 福州大學環境與資源學院, 福州 350116
植物功能性狀(plant function traits,PFT)是指影響植物個體生存、發育和繁殖所表現出的生理、形態和生活習性上的特性[1-2]。它是植物長期演化過程中對不同生活環境適應的結果[3],反映了植物對生存環境中各種資源的利用情況與適應性[4-6],是連接植物與環境的橋梁[7]。葉片是植物進行光合作用,合成光合產物以及獲取養分的重要場所,它與環境接觸面積最大、是植物對環境變化響應明顯的敏感性器官[5]。在植物各器官功能性狀中,葉功能性狀(leaf functional traits,LFT)與植物的生存策略及資源利用能力等聯系密切[8],能夠直接反映植物適應環境所形成的生存對策[9- 11]。葉功能性狀可以劃分為兩大類型,即葉結構型性狀和葉功能型性狀。其中前者是指植物葉片的生物化學結構特征,它在一定環境中保持相對穩定不變,并且能反映植物為獲得最大程度的碳收入而采取的適應性策略[12];后者則是反映葉片生長代謝的指標,它會隨著時間和空間的變化而改變,穩定性相對較差,難以深入解釋植物在長期進化過程中的適應策略[13]。因此,本文僅從葉結構型性狀方面開展亞熱帶常綠闊葉林主要樹種對毛竹擴張的響應與適應機制研究,具有重要的生態學意義。
毛竹(Phyllostachysedulis)屬于禾本科(Poaceae)竹亞科(Bambusoideae)剛竹屬(Phyllostachys),是一種特殊的高大克隆喬木狀散生植物。它依靠其鞭根進行無性繁殖,向臨近的林分內生長并逐漸擴張、蔓延,具有強大的繁衍生殖和空間拓展能力[14]。亞熱帶常綠闊葉林是我國重要的地帶性植被,具有豐富的生物多樣性資源和巨大的生態效益[15],在調節氣候、涵養水源和保持水土等方面發揮著重要作用[16]。由于毛竹在向臨近常綠闊葉林擴張過程中,與闊葉樹種的生態位存在著交叉或重疊[16],其必然會與闊葉樹種爭奪光、水、熱、土壤養分以及地上空間等資源,可能導致林木葉片功能性狀發生變化,進而影響林木的正常生長與生存。目前,有關毛竹擴張對亞熱帶常綠闊葉林影響的研究多集中于毛竹擴張對森林的物種組成[17]、群落結構[18]、生物多樣性[16]、N循環[19]等群落學與生態效應影響方面,而有關毛竹擴張對常綠闊葉林林木葉功能性狀影響方面的研究較少。本文以存在于常綠闊葉林和竹闊混交林中的四個相對優勢樹種,即常綠闊葉樹種甜櫧(Castanopsiseyrei)和青岡(Cyclobalanopsisglauca)、落葉闊葉樹種青榨槭(Acerdavidii)和枳椇(Hoveniaacerba)為研究對象,試圖探究:(1)毛竹擴張是否會改變喬木層林木葉結構型性狀特征?(2)毛竹擴張對不同生活型林木葉結構型性狀的影響是否具有一致性?(3)毛竹擴張是否改變喬木層4種林木間葉結構型性狀的差異特征?(4)毛竹擴張是否會影響喬木層林木葉結構型性狀指標間的關系特征?研究結果可為探明我國亞熱帶常綠闊葉林主要森林植物葉功能性狀對毛竹擴張的響應機制,進一步開展毛竹擴張對常綠闊葉林的林分質量與生態功能的影響機制研究等提供理論依據,并為適度控制毛竹擴張、開展常綠闊葉林生態系統健康研究等提供實踐指導。
1 材料與方法
1.1 研究區概況
江西陽際峰自然保護區地處江西省東北部,貴溪市境內(27°51′07″—28°00′24″N, 117°13′42″—117°24′22″E),屬武夷山脈中段西北坡(圖1),保護區面積10946 hm2,森林覆蓋率高達99.7%,境內保存有6000余hm2原生性較強的常綠闊葉林,1878種高等植物,分別屬8個植被型72個群系,240科830屬,是我國亞熱帶常綠闊葉林的典型代表。保護區屬中亞熱帶濕潤季風山地氣候,年平均氣溫11.4—18.5℃,年平均降水量1870—2191.3 mm,無霜期200—256 d,區內氣象災害少,光、溫、水等氣候資源時空分布較均勻,有利于動植物生長及繁衍,為科學研究提供了理想的研究平臺。

圖1 研究區位置圖
1.2 樣地設置與樣品采集
在江西省陽際峰國家級自然保護區內,沿毛竹水平擴張方向,分別設置6個常綠闊葉林樣地與6個竹闊混交林樣地,共12個樣地。樣地面積為15 m×15 m。所選取的樣地海拔在700—1200 m,竹闊混交林樣地受毛竹擴張影響達20余年。于生長季(2017年6月)對樣地內毛竹及胸徑5.0 cm以上的林木進行每木調查,并分樹種計算其平均胸徑、平均高及重要值。選取在常綠闊葉林與竹闊混交林喬木層中都有分布且具有一定代表性的兩常綠闊葉樹種(甜櫧與青岡)與兩落葉闊葉樹種(青榨槭與枳椇)共4樹種,對其葉結構型性狀進行分析。4樹種重要值分別為甜櫧14.68%、青岡10.99%、青榨槭12.18%、枳椇3.16%,為陽際峰自然保護區常綠闊葉林中2生活型植物(常綠闊葉與落葉闊葉)的相對優勢種。對所選取的4樹種分別樣地、分樹種選擇胸徑、株高接近其平均胸徑和平均高的健康木3株進行葉片采樣。選取林冠中層完全展開的外層成熟健康葉片進行采樣,所采葉片的葉齡均為當年生葉,分東、南、西、北四個方向選取1小枝,每個枝條選3—5片葉。將所采集葉片立即置于塑封袋中,編號帶回實驗室。樣地基本情況見表1與表2。
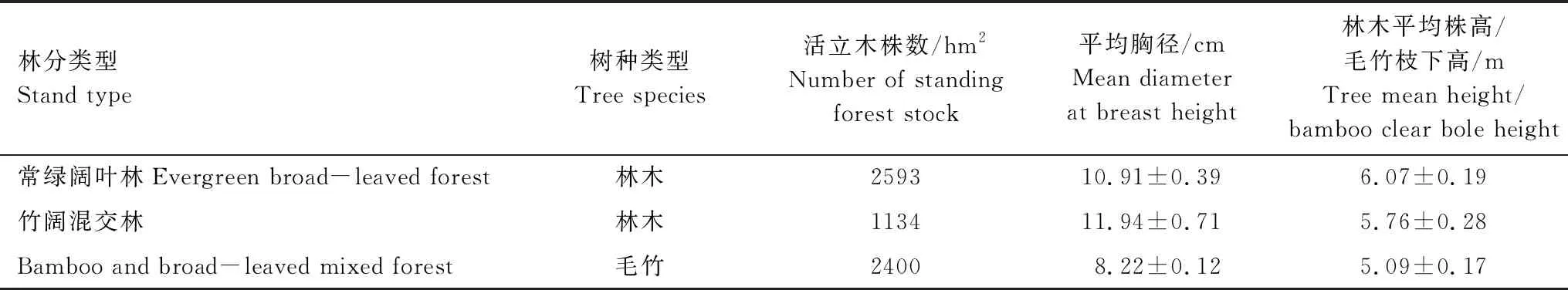
表1 江西陽際峰國家級自然保護區不同林分樣地概況
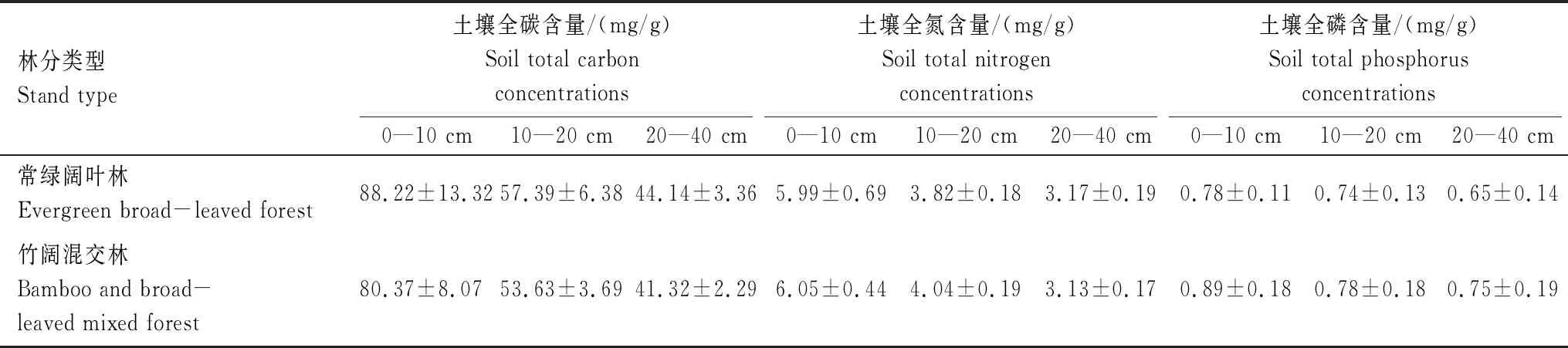
表2 江西陽際峰國家級自然保護區不同林分樣地土壤養分含量
1.3 葉功能性狀指標選取與測定
選取葉長(leaf length, LL)、葉寬(leaf width, LW)、葉面積(leaf area, LA)、比葉面積(specific leaf area, SLA)、葉厚(leaf thickness, LT)、葉鮮重(leaf fresh weight,LFW)、葉干重(leaf dry weight, LDW)、葉干物質含量(leaf dry matter content, LDMC)、葉組織密度(leaf tissue density, LTD)、葉相對含水量(leaf relative water content, LRWC)、葉形態指數(leaf morphology index, LMI)等11個葉結構型性狀進行分析。葉結構型性狀測定步驟如下:
利用EPSON V19掃描儀掃描葉片,用Image J軟件計算LL、LW、LA;用游標卡尺(精度0.01 mm)測定葉片沿著主脈同一側方向上、中、下3個位置上的厚度(避開主葉脈),取平均值即為該葉片LT;采用電子天平(精度0.01 g)稱取LFW后,將葉片樣品浸泡于清水中,于5℃的黑暗環境中浸12 h,取出后迅速用吸水紙去除葉片表面的水分,稱其飽和鮮重;之后將其放在75℃下恒溫箱內烘干48—72 h后稱重,稱LDW。計算其LRWC、LDMC、SLA、LMI及LTD等葉性狀指標。計算公式如下:
SLA(cm2/g)=葉面積(cm2)/葉干重(g)
LDMC(g/kg)=葉干重(g)/葉飽和鮮重(g)
LRWC(%)=[葉鮮重(g)-葉干重(g)]/[葉飽和鮮重(g)-葉干重(g)]×100%
LTD(g/cm3)=葉干重(g)/[葉面積(cm2)·葉厚(mm)]
LMI(cm/cm)=葉長(cm)/葉寬(cm)
1.4 數據處理
利用Canoco 5軟件中的主成分分析法分析并篩選出對各樹種具有重要影響的主要葉結構型性狀指標;據此,運用SPSS20.0軟件中的獨立樣本T檢驗法分析毛竹擴張對4喬木層林木總體及不同生活型樹種主要葉結構型性狀影響,并采用雙因素方差分析法分析毛竹擴張對各樹種主要葉結構型性狀的影響。利用模型lgY=lgα+βlgM(式中Y、M分別代表兩主要葉結構型性狀指標,α為截距,β為斜率)模擬竹闊混交林與常綠闊葉林內林木主要葉結構型性狀間關系,分析毛竹擴張對其主要葉結構型性狀關系的影響。利用R3.5.2數據分析軟件中的標準化主軸回歸分析法(SMA)計算所選模型的截距、斜率及斜率的置信區間,并對斜率進行異質性檢驗,在斜率同質時計算其共同斜率[20]。
2 結果與分析
2.1 林木主要葉結構型性狀指標的篩選
為了更好地揭示各樹種葉結構型性狀的內在規律,對所測定的LL、LW、LA、SLA、LT、LFW、LDW、LDMC、LTD、LRWC以及LMI等11個葉結構型性狀進行主成分分析(PCA)。結果顯示:第1主成分(第1排序軸)解釋了51.52%的變異,第2主成分(第2排序軸)解釋了21.31%的變異,累計解釋量為72.83%。由此可見,前兩個主成分能較好地反映對樹種影響較大的葉結構型性狀,且其影響大小主要由第一主成分決定。其中與第一主成分相關最密切的葉結構型性狀指標主要為LL、LW、LA、SLA、LTD、LMI及LDMC,且LL、LW、LA和SLA均與第一主成分軸呈正相關,LTD、LMI和LDMC則均與第一主成分軸呈負相關,其相關性大小依次為:LW>LA> LL> LDMC >LTD >SLA >LMI(圖2)。因此,本研究確定選取LL、LW、LA、SLA、LTD、LMI和LDMC 共7個指標作為主要葉結構型性狀,用于分析毛竹擴張對林木葉結構型性狀的影響。
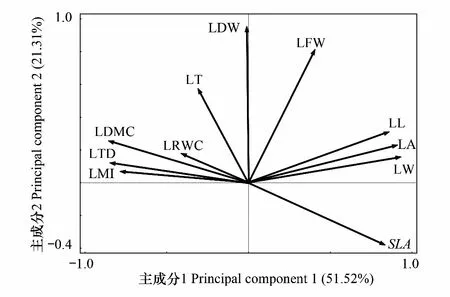
圖2 毛竹擴張下4種主要植物葉結構型性狀主成分分析圖
2.2 毛竹擴張對喬木層林木總體葉結構型性狀的影響
以闊葉林中的甜櫧、青岡、青榨槭、枳椇4主要樹種的各葉結構型性狀均值進行分析,結果發現,4林木的LL、LW、LA、LTD、LMI、LDMC及SLA均值在常綠闊葉林與竹闊混交林之間雖存在差異,但差異不明顯(P>0.05)(圖3),說明毛竹擴張對常綠闊葉林喬木層林木總體葉結構型性狀的影響不明顯。
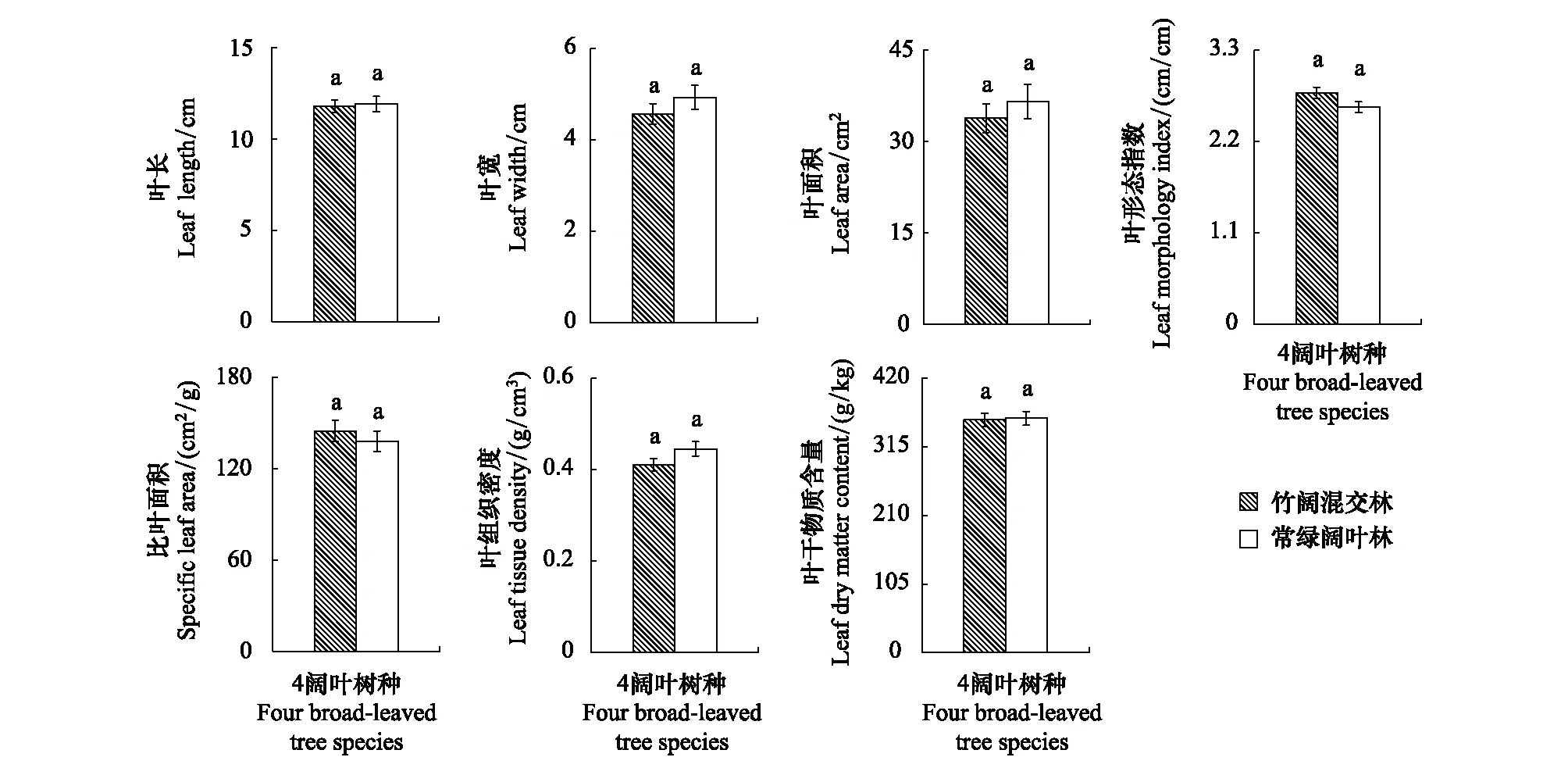
圖3 喬木層林木同一葉結構型性狀指標在不同林分的表現特征
2.3 毛竹擴張對不同生活型闊葉樹種葉結構型性狀的影響
通過對兩常綠闊葉樹種與兩落葉闊葉樹種的主要葉結構型性狀分析,結果顯示,毛竹擴張對常綠闊葉林喬木層不同生活型闊葉樹種葉結構型性狀的影響存在差異(圖4),常綠闊葉林內落葉闊葉樹種葉結構型性狀對毛竹擴張影響的敏感性相對高于常綠闊葉樹種。毛竹擴張對常綠闊葉樹種的LTD及落葉闊葉樹種的LW與LMI兩葉結構型性狀指標具有顯著影響(P<0.05)。竹闊混交林中常綠闊葉樹種的LTD顯著小于其在常綠闊葉林的LTD(P<0.05),而其LL、LW、LA、SLA、LDMC以及LMI等葉結構型性狀指標在竹闊混交林與常綠闊葉林兩林分內則差異不顯著(P>0.05)。竹闊混交林中落葉闊葉樹種的LMI顯著高于常綠闊葉林(P<0.05),但其LW則顯著小于常綠闊葉林(P<0.05),而其LL、LA、SLA、LTD以及LDMC等葉結構型性狀指標在兩林分內差異不顯著(P>0.05)。
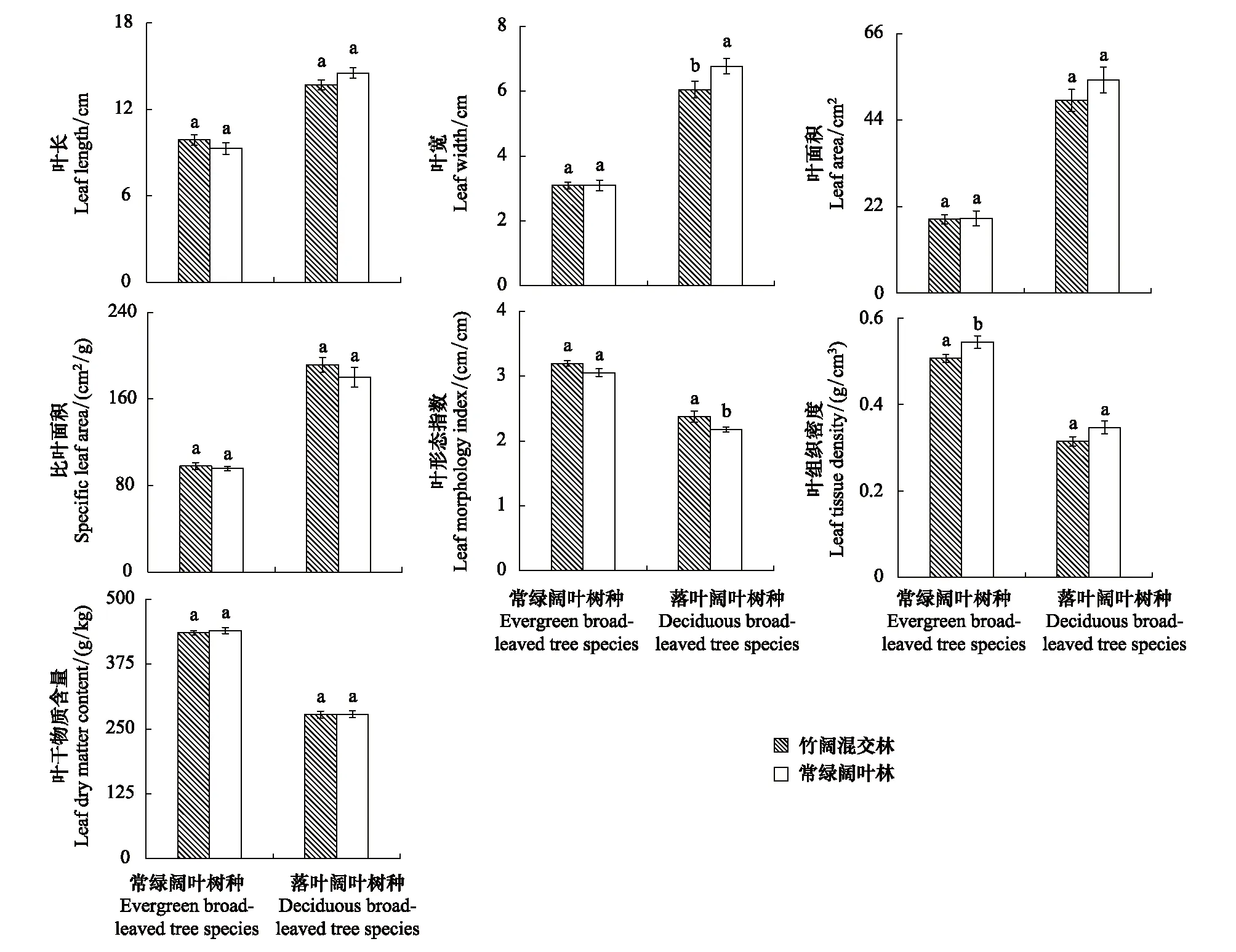
圖4 不同生活型樹種葉結構型性狀指標的比較
2.4 毛竹擴張對不同樹種葉結構型性狀的影響
不同樹種對毛竹擴張影響的響應程度不同(圖5)。竹闊混交林中的甜櫧LL、LW和LA顯著大于常綠闊葉林(P<0.05),但其LMI、SLA、LTD和LDMC在兩林分間則差異不顯著(P>0.05);毛竹擴張會使竹闊混交林中青岡的LMI顯著增加,明顯高于常綠闊葉林的LMI(P<0.05),但其LTD、LDMC則顯著小于常綠闊葉林(P<0.05),而其LL、LW、LA和SLA在兩林分間則差異不顯著(P>0.05);竹闊混交林中的枳椇LMI顯著高于常綠闊葉林(P<0.05),但其LW與LTD則顯著小于常綠闊葉林(P<0.05),而其LL、LDMC、LA和SLA在兩林分間則差異不顯著(P>0.05);竹闊混交林中的青榨槭LW顯著小于其在常綠闊葉林的LW(P<0.05),但其在兩林分內的LL、LA、SLA、LMI、LDMC和LTD則差異不顯著(P>0.05)。由此可見,隨著毛竹向常綠闊葉林的擴張,4喬木層林木的部分葉結構型性狀會發生顯著的改變。
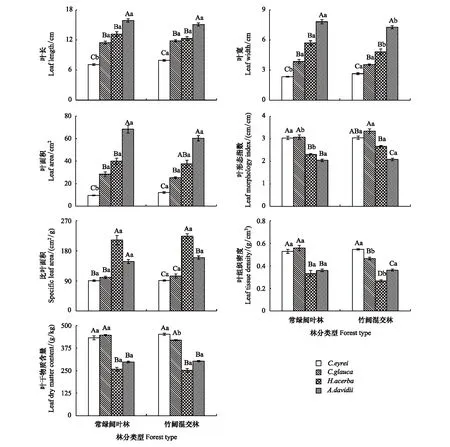
圖5 不同樹種葉結構型性狀指標的比較
毛竹擴張還會改變不同樹種間部分葉結構型性狀的差異特征(圖5)。毛竹擴張會改變喬木層4種林木間LTD與LDMC兩性狀指標的大小次序。4林木LTD大小次序在常綠闊葉林中其表現為:青岡>甜櫧>青榨槭>枳椇,其中,甜櫧和青岡的LTD顯著大于枳椇及青榨槭(P<0.05),且甜櫧和青岡兩常綠闊葉樹種間及枳椇和青榨槭兩落葉闊葉樹種間的LTD差異均表現不明顯(P>0.05);但在竹闊混交林中其LTD大小次序則表現為:甜櫧>青岡>青榨槭>枳椇,且各樹種間LTD均存在顯著差異(P<0.05)。喬木層4林木間LDMC在常綠闊葉林中的大小次序為:青岡>甜櫧>青榨槭>枳椇,但在竹闊混交林中其大小次序則表現為:甜櫧>青岡>青榨槭>枳椇;兩森林類型均表現為:青岡和甜櫧兩常綠闊葉樹種的LDMC顯著大于枳椇和青榨槭兩落葉闊葉樹種(P<0.05),但兩常綠闊葉樹種間及兩落葉闊葉樹種間的LDMC差異均不明顯(P>0.05)。4林木間的LMI與SLA雖在常綠闊葉林與竹闊混交林中的大小次序未發生改變,但各樹種間差異的顯著性則在兩森林類型中表現不同。4林木間的LL、LW與LA在常綠闊葉林與竹闊混交林中的大小次序及其差異顯著性均表現為一致。上述結果說明,毛竹擴張會改變喬木層4林木間LMI、LTD、SLA及LDMC的差異特征,但不會改變4林木間的LL、LW與LA差異特征。
2.5 毛竹擴張對喬木層林木葉結構型性狀間關系影響分析
毛竹擴張會影響常綠闊葉林喬木層林木部分葉結構型性狀間關系(圖6)。LL與LW、LW與LA在竹闊混交林與常綠闊葉林中雖然均分別擁有共同斜率,但竹闊混交林中LL與LW、LW與LA的截距均顯著小于常綠闊葉林,這說明毛竹擴張會改變常綠闊葉林內喬木層林木的LL與LW、LW與LA之間的數量關系。但毛竹擴張對LA與LL、SLA與LA、SLA與LTD、SLA與LDMC間的關系影響較小,它們在竹闊混交林與常綠闊葉林中擁有共同斜率(即異速生長指數)及共同截距(即異速生長常數)(圖6),說明毛竹擴張并不會改變常綠闊葉林內喬木層林木LA與LL、SLA與LTD、SLA與LA、SLA與LDMC性狀間關系。
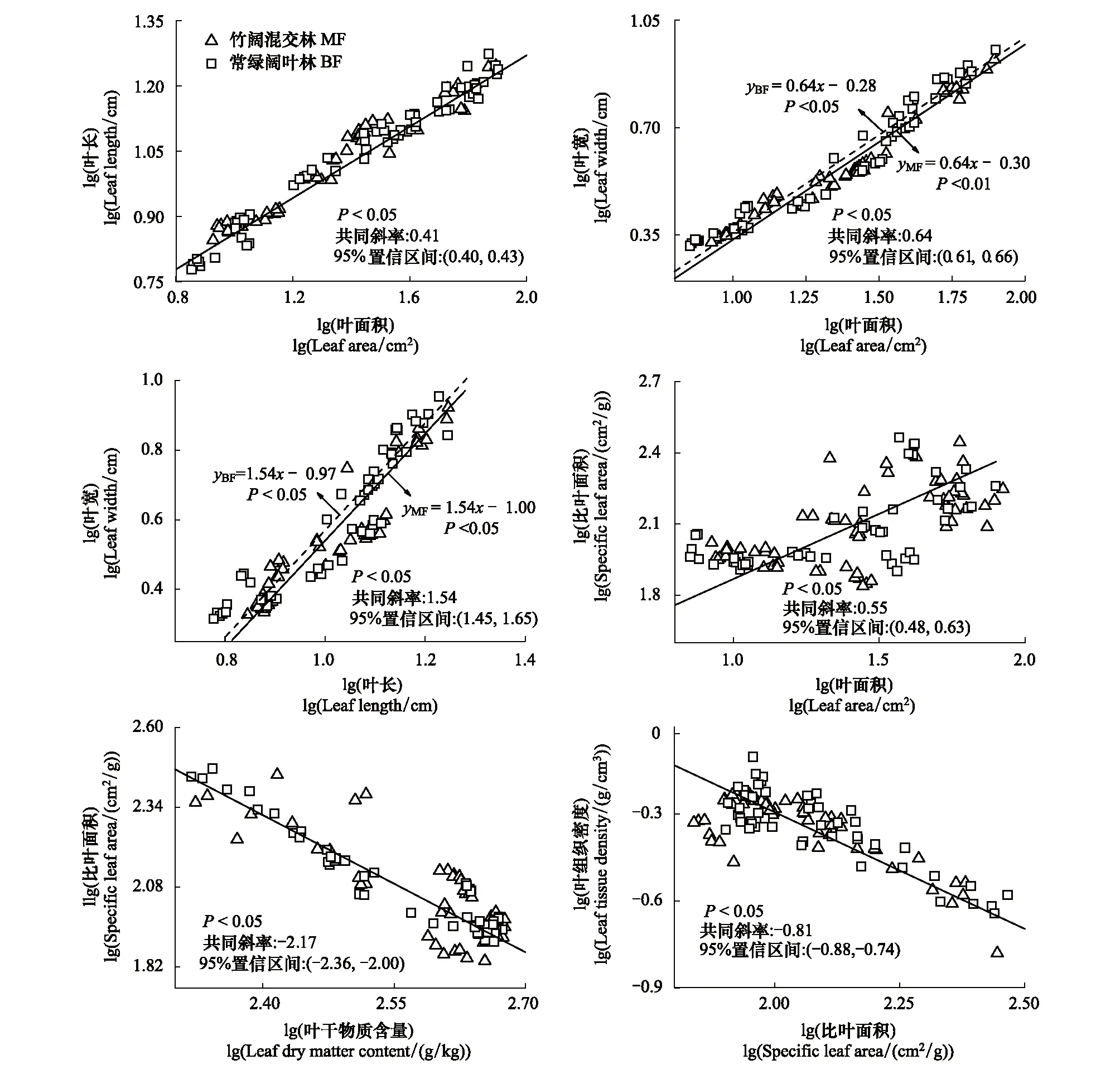
圖6 不同林分內林木LL、LW、LA、LTD、LT、SLA之間的線性關系
3 討論
3.1 毛竹擴張對喬木層林木總體葉結構型性狀特征的影響
植物葉性狀的分異主要受植物類群、植物體高度和生活型影響,植物類群對植物葉性狀的影響要大于環境因子(如溫度、水分和經緯度等),以科類群影響為最大[21- 22]。本研究結果顯示,4喬木層樹種總體葉結構型性狀特征在竹闊混交林與常綠闊葉林雖存在差異,但差異不顯著,分析原因,主要是由于本研究所選擇的樹種是兩森林類型內共有種,其葉性狀主要受植物種影響,毛竹擴張后,雖然會對森林內植物的光照、水分等環境因子造成影響,但環境對葉性狀的影響要小于植物種影響,從而導致毛竹擴張對常綠闊葉林喬木層林木總體葉結構型性狀影響不明顯。
3.2 毛竹擴張對不同生活型闊葉樹種葉結構型性狀的影響
在森林生態系統中,不同生活型植物的表型可塑性不同。有研究顯示,常綠闊葉植物與落葉闊葉植物在SLA、葉片壽命、礦質元素含量等葉性狀方面存在差異[23],而常綠闊葉樹種在競爭激烈的環境中往往比落葉闊葉樹種更占優勢[24]。受毛竹擴張影響,常綠闊葉林中的常綠闊葉樹種僅有LTD發生顯著變化,毛竹擴張會導致常綠闊葉樹種LTD顯著降低;而落葉闊葉樹種則表現為LW與LMI兩葉結構型性狀指標變化明顯,毛竹擴張會導致落葉闊葉樹種的LW顯著減小、LMI則顯著升高,反映了落葉闊葉樹種對毛竹擴張的敏感程度相對高于常綠闊葉樹種。有研究表明, LTD與植物的碳儲備能力有關,LTD較高體現了葉片以慢速周轉方式生長,具有較多的碳儲備[25]。毛竹擴張會使常綠闊葉林中的常綠闊葉樹種LTD顯著減小,反映了常綠闊葉樹種為適應毛竹擴張而采取開拓性生長策略,即植物主要投資于生長速率和資源獲取能力[26],減少碳儲備而將更多養分投入到高生長以保持自身的競爭優勢。受毛竹擴張影響,闊葉林中的落葉闊葉樹種則通過減小LW,增大LMI,調整葉片形態,減少對保護組織的投資,調節葉片對散射光與漫射光的吸收,增大其耐陰性,以提高其對毛竹擴張造成光資源相對不足的環境適應能力[27]。
3.3 毛竹擴張對4喬木層林木葉結構型性狀的影響
植物葉片對環境變化敏感且可塑性大[28],可以反映植物對環境的適應能力[8]。不同樹種間同一葉結構型性狀指標對毛竹擴張影響的響應程度存在差異,同一樹種各葉結構型性狀指標對毛竹擴張影響的響應程度也不同。甜櫧是本研究區域常綠闊葉林中的優勢種群,屬于殼斗科(Fagaceae),錐屬(Castanopsis),具有較高的耐蔭性與資源爭奪能力[29],毛竹擴張會使甜櫧LL、LW以及LA顯著增大,即表現為葉片由小到大的變化趨勢,反映了甜櫧對毛竹擴張后光環境減弱的適應性,增大其光合作用面積,補償較低光量子通量密度造成的光合速率下降,從而保持其資源競爭優勢[30],以促進其生存與生長。青岡為殼斗科青岡屬(Cyclobalanopsis),耐干旱瘠薄,是適應能力強的廣適種,面對毛竹擴張,其LTD和LDMC顯著減小,表明青岡為適應毛竹擴張所引起的養分及水熱資源爭奪激烈的環境,減少養分的儲備以及對結構組織構建的投資,從而將更多的養分用于周轉及生長;而其LMI顯著增大,葉片從橢圓形向細長型變化,體現了葉片形態對光脅迫的適應。甜櫧與青岡雖同屬于殼斗科植物,但受毛竹擴張影響,甜櫧LL、LW及LA發生顯著改變,而青岡的LL、LW及LA則變化不明顯,但其LTD、LDMC、LMI則發生了顯著改變,這表明即使是同科植物面對毛竹擴張其適應策略也不相同。枳椇為鼠李科(Rhamnales)枳椇屬(Hovenia)植物,生長快、易存活、對土壤氣候要求不高,毛竹擴張會導致其LW與LTD顯著減小、LMI顯著增大,說明該樹種受毛竹擴張影響,會通過改變葉形態,減小碳存儲投資,提高養分運轉利用效率以獲得競爭優勢,從而適應毛竹擴張對其生長的影響。青榨槭為槭樹科(Aceraceae)槭屬(Acer)喬木,形態可塑性高,適應性強,既可在溝谷、林緣等光源充足的開闊地生長,又可在光源競爭激烈的林下生長[31],毛竹擴張僅會造成其LW顯著減小,說明該樹種的葉結構型性狀受毛竹擴張影響較小。
3.4 毛竹擴張對喬木層林木葉結構型性狀間關系的影響
植物為了適應環境的變化,通過權衡資源分配和調節生理過程,在形態上表現出特殊的植物功能性狀的變化[32-33],進而形成一系列性狀組合[34-35],具有一定的可塑性。毛竹擴張會對喬木層林木部分葉結構型性狀間關系產生顯著影響,使喬木層林木LL與LW、LW與LA間的截距顯著減小,表明喬木層林木面對毛竹擴張,在相同的LL投入下,竹闊混交林中喬木層林木LW的投入相對較小;而在相同的LA投入下,竹闊混交林中喬木層林木LW的投入也相對較小。這主要是由于毛竹擴張加劇植物對光、水、熱等資源的競爭,而水脅迫會影響葉片的形態。有研究顯示,LW減小有利于植物葉片的直接散熱,而不通過蒸騰降溫[36],防止水分流失,提高喬木層林木對毛竹擴張造成的林地土壤水分降低壓力的適應能力。
毛竹擴張可能還會對常綠闊葉林內喬木層其他林木的葉功能型性狀產生影響,并可能會對莖干與細根功能性狀產生影響,這些研究有助于探清常綠闊葉林主要樹種對毛竹擴張的響應與適應機制。
4 結論
毛竹擴張對常綠闊葉林喬木層4樹種總體葉結構型性狀無明顯影響,但其對不同生活型闊葉樹種葉結構型性狀的影響存在差異,落葉闊葉樹種葉結構型性狀對毛竹擴張影響的敏感性相對高于常綠闊葉樹種。毛竹擴張會造成喬木層樹種的部分葉結構型性狀發生顯著改變。受毛竹擴張影響,同一葉結構型性狀變化在不同樹種間存在差異;同一樹種不同葉結構型性狀對毛竹擴張的響應程度也存在差異。毛竹擴張會導致常綠闊葉林喬木層樹種部分葉結構型性狀間關系發生改變,它會改變林木LL與LW、LW與LA之間的數量關系。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
