乙烯信號傳導途徑因子OsEIL6調控水稻抗稻瘟病反應
2020-08-25 10:03:25趙玉丹朱曉穎徐國娟王旭麗
植物保護 2020年4期
關鍵詞:水稻
趙玉丹 朱曉穎 徐國娟 王旭麗
摘要 稻瘟病(rice blast)是水稻生產上最嚴重的病害之一。抗病相關基因的挖掘對稻瘟病的防治具有重要意義。研究表明植物EIN3/EIL家族基因在抗病過程中發揮著重要作用。本研究采用RNAi技術探究OsEIL6參與的水稻抗稻瘟病反應。稻瘟菌侵染時基因表達譜檢測結果表明,OsEIL6在水稻和稻瘟菌非親和組合中受到誘導表達。稻瘟菌接種結果顯示,水稻OsEIL6沉默株系和野生型植株‘TG394相比抗性下降;實時熒光定量RT-PCR結果分析表明,OsEIL6的表達量下降導致乙烯合成途徑中OsACO1和乙烯信號傳導途徑的OsERF063和OsERF073的轉錄水平下降。亞細胞定位研究發現該基因定位于水稻細胞質。OsEIL6沉默株系中ROS合成途徑標記基因OsrbohA和OsrbohB的表達量均明顯下調,表明該基因可能通過影響ROS的合成調控水稻抗稻瘟病反應。本研究結果將有助于進一步揭示OsEIL6參與的乙烯信號傳導途徑介導的水稻抗稻瘟病反應機制。
關鍵詞 水稻; 稻瘟病; 抗病性; 乙烯; OsEIL6
中圖分類號: S 432.1
S 435.121.45
文獻標識碼: A
DOI: 10.16688/j.zwbh.2020153
OsEIL6 is involved in regulating rice resistance to Magnaporthe oryzae
ZHAO Yudan#, ZHU Xiaoying#, XU Guojuan, WANG Xuli*
(State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection,
Chinese Academy of Agricultural Sciences, Beijing 100193, China)
Abstract
Rice blast is one of the most destructive rice diseases. Understanding the function of rice immune responsive genes to the blast fungus Magnaporthe oryzae is essential for effective disease management. Previous research revealed that ethylene-insensitive 3 (EIN3) /ethylene-insensitive 3-like (EIL) gene plays essential roles in plant defense responses. In this study, we investigated the role of OsEIL6 in rice resistance to M.oryzae. Expression of OsEIL6 was significantly induced in rice after infection with an incompatible blast isolate C9240. Silencing of OsEIL6 in transgenic rice enhanced plant susceptibility to M.oryzae. Quantitative RT-PCR assays showed that the transcript levels of OsACO1 in ethylene biosynthesis, and two key downstream genes, OsERF063 and OsERF073, in ethylene signaling pathways, ware down-regulated in the OsEIL6-RNAi plants. Subcellular localization analyses showed that the OsEIL6 protein was localized in the cytoplasm of rice cells. In addition, the expression levels of OsrbohA and OsrbohB, two key genes for ROS biosynthesis, were down-regulated in the OsEIL6-RNAi plants, suggesting that OsEIL6 may mediate rice resistance by affecting the ROS production. Our findings will contribute to further understanding of the mechanism of OsEIL6-mediated resistance to M.oryzae.
Key words
Oryza sativa; rice blast; disease resistance; ethylene; OsEIL6
水稻是最重要的糧食作物之一,世界上有超過一半的人口以其為主糧[1]。因此,水稻的豐產、穩產對糧食安全具有極其重要的戰略意義。水稻稻瘟病危害重、防治難,已成為影響水稻豐產穩產的關鍵因素。據統計,全球由于稻瘟病危害造成的產量損失高達水稻年總產的10%~30%[2-3]。目前,防治稻瘟病最為經濟有效和環保的措施是種植抗病品種,而抗病品種的培育則依賴于全面深入認識水稻抗病機理。
為了抵御病原物的入侵,植物進化出兩個層次的免疫反應PTI(PAMP-triggered immunity)和ETI(effector-triggered immunity)[4]。而在PTI和ETI所激活的下游免疫反應中,激素信號途徑發揮著重要作用,其中乙烯(ethylene, ET)信號扮演重要角色[1, 4]。乙烯是一種小分子氣體植物激素,參與調控植物生命活動的許多過程,如促進葉片和果實成熟,以及植物對生物和逆境脅迫的反應等。在模式植物擬南芥中,結合遺傳學、生物化學和分子生物學等技術,科學家們發現了一條近乎線性的乙烯信號傳導途徑[5]。近年來,在水稻中也相繼鑒定了乙烯信號傳導的關鍵因子,解析了水稻乙烯信號傳導途徑[6]。研究表明組氨酸激酶(histidine kinase)家族類受體蛋白識別乙烯后可失去對OsCTR1的抑制,隨后觸發轉錄因子EIN2(ethylene insensitive 2)及其下游EIN3/EIL(ethylene insensitive 3/EIN3-like)介導的級聯反應[7]。最近,Zhao等在水稻中鑒定了一個乙烯信號傳導的新組分—MHZ1/OsHK1,其介導一條平行于OsEIN2-OsEIL1通路的乙烯信號傳導新途徑,但乙烯對MHZ1轉錄的促進仍然依賴于OsEIL1[8]。綜上,EIN3/EIL家族基因處于乙烯信號傳導的中樞位置,決定著乙烯下游響應基因的表達。
EIN3/EIL家族基因在調控植物的抗病反應中發揮著重要作用[9-11]。在擬南芥中EIN3和EIL1可抑制SA合成基因ICS/SID2的表達,導致SA合成量減少,負調控擬南芥對丁香假單胞菌Pseudomonas syringae的抗性[9]。其次,EIN3和EIL1可通過與JA信號通路的MYC2蛋白相互作用,整合ET和JA信號,正調控擬南芥對灰葡萄孢Botrytis cinerea的抗性[10]。水稻該家族基因OsEIL1表達量的下降可引起乙烯含量的下降和對稻瘟病抗性的減弱,OsEIL1通過靶標ROS和茉莉酸(jasmonate, JA)合成的關鍵酶基因來介導其正向調控水稻抗稻瘟病反應[11]。由此可見,EIN3/EIL家族基因既可正調控又可負調控植物抗病反應,其作用結果與病菌和寄主間相互作用的具體特點有關。水稻基因組EIN3/EIL家族共有6個基因OsEIL1-6[12-13],本實驗室報道了OsEIL2參與調控水稻抗紋枯病反應[14],進一步檢測發現OsEIL6的表達受稻瘟菌誘導。本研究將探究OsEIL6參與調控的水稻抗稻瘟病反應,研究結果將有助于闡明水稻中乙烯信號途徑所參與的抗病反應過程,并可為水稻稻瘟病防控提供新思路。
1 材料與方法
1.1 材料與試劑
水稻品種‘日本晴以及稻瘟菌親和小種RO1-1和非親和小種C9240為本實驗室保存,轉化用水稻品種‘泰粳394(‘TG394)為揚州大學左示敏老師惠贈。試驗所用亞細胞定位載體pYBA-1132和蛋白表達載體pRTV-nHA均為本實驗室保存。引物合成和測序由北京擎科生物公司完成。
1.2 試驗方法
1.2.1 載體構建
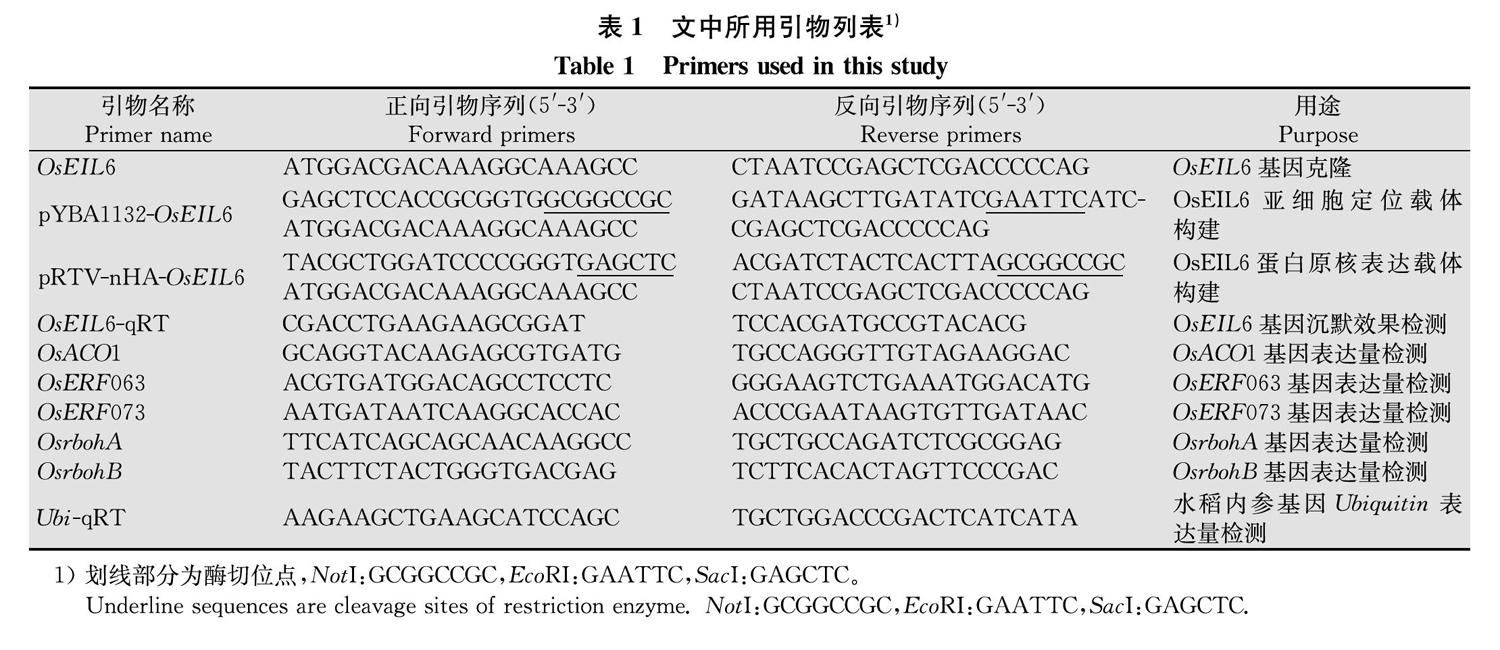
根據MSU(http:∥rice.plantbiology.msu.edu/)數據庫中的OsEIL6基因序列,設計特異性引物OsEIL6-F和OsEIL6-R(表1)擴增獲得OsEIL6基因全長。
為了確定OsEIL6的亞細胞定位,設計特異性引物pYBA1132-OsEIL6-F和pYBA1132-OsEIL6-R(表1)擴增獲得帶有酶切位點的片段,采用同源重組的方法將擴增得到的片段連接到融合GFP標簽的表達載體pYBA1132上,獲得亞細胞定位表達載體pYBA1132-OsEIL6。
為了表達和純化OsEIL6蛋白,用特異性引物pRTV-nHA-OsEIL6-F和pRTV-nHA-OsEIL6-R(表1)擴增獲得帶有酶切位點的片段,同源重組連接到載體pRTV-nHA上,獲得蛋白表達載體pRTV-nHA-OsEIL6。
1.2.2 水稻轉基因材料的單拷貝純合株系的獲得
按照Park等[15]的試驗方法,將供試的水稻材料種子經表面消毒后放置于1/2 MS培養基上進行催芽。在T2代培養時,選取陽性株和陰性株比例為3∶1的轉基因株系,鋪種于含有潮霉素的1/2 MS培養基上生長,篩選出陽性率為100%的轉基因株系即為單拷貝純合株系,用于后續的接種試驗。
1.2.3 稻瘟菌接種與病情調查
參照Yang等[11]的方法,并略作改動。選用培養3周的水稻苗用于接種。將配制好的濃度為1.2×105個/mL的供試菌株孢子懸浮液均勻地噴灑在葉片表面,26~28℃條件下黑暗保濕培養24 h后,轉入光照條件下繼續培養,7 d左右調查病情。采用Photoshop軟件對所有拍照病葉進行病斑面積計算,并通過GraphPad Prism軟件對數據進行分析作圖,進行多重比較和單因素方差分析。
1.2.4 RNA提取、反轉錄及qRT-PCR分析
采用Trizol法提取水稻葉片總RNA,用DNase I去除DNA后用反轉錄試劑盒(Promega)反轉錄為cDNA。選用ABI-7500型熒光定量PCR儀進行定量檢測。qRT-PCR的體系為:2×SYBR Premix ExTaq 10 μL,稀釋后的cDNA 5 μL,正反向引物各1 μL,最后用ddH2O補齊至20 μL。qRT-PCR的程序為:95℃ 30 s;95℃ 15 s,55℃ 30 s,72℃ 30 s,反應共進行48個循環;繼續進行熔解曲線反應(55℃,81個循環),檢測產物的單一性。以Ubiquitin作為內參基因,根據相對閾值法(2-ΔCT)計算基因的表達量,每個樣品做3次重復,數據選取3次重復的平均值,通過GraphPad Prism軟件進行多重比較和單因素方差分析。
1.2.5 水稻原生質體的制備與轉化
‘日本晴水稻種子經75%乙醇浸泡1 min,40%次氯酸鈉 浸泡30 min,無菌水清洗3遍后,播種在1/2 MS培養基上,放置在26~28℃培養箱黑暗培養10 d左右。按照Zhang等[16]原生質體提取和亞細胞定位的方法,將獲得的水稻黃化苗莖稈切成長度為0.5 mm左右的莖段,隨后轉移至酶解液(1.0%纖維素酶RS,0.5%離析酶R10,0.6 mol/L甘露醇,10 mmol/L MES,pH調至5.7,10 mmol/L CaCl2和0.1% BSA)中28℃酶解5~7 h,40 μm尼龍篩過濾收集原生質體,利用PEG介導轉化法將目的質粒轉化到原生質體中,26~28℃培養箱中培養18~24 h后,激光共聚焦顯微鏡下觀察拍照。
1.2.6 液相色譜-質譜聯用 (liquid chromatography-mass spectrometry, LC-MS)方法
利用PEG介導轉化法將目的質粒pRTV-nHA-OsEIL6轉化到水稻原生質體中,在原生質體中表達帶有HA標簽的OsEIL6蛋白,26~28℃培養箱中培養16~24 h后,利用非變性蛋白提取液提取水稻原生質體蛋白,利用免疫沉淀(IP)技術富集帶有HA標簽的OsEIL6蛋白,蛋白凝膠電泳5 min后回收膠條,送至中科新生命科技有限公司進行液相色譜-質譜聯用(LC-MS)分析,鑒定混合物中的蛋白。
2 結果與分析
2.1 水稻OsEIL6在稻瘟病菌非親和小種侵染時誘導表達
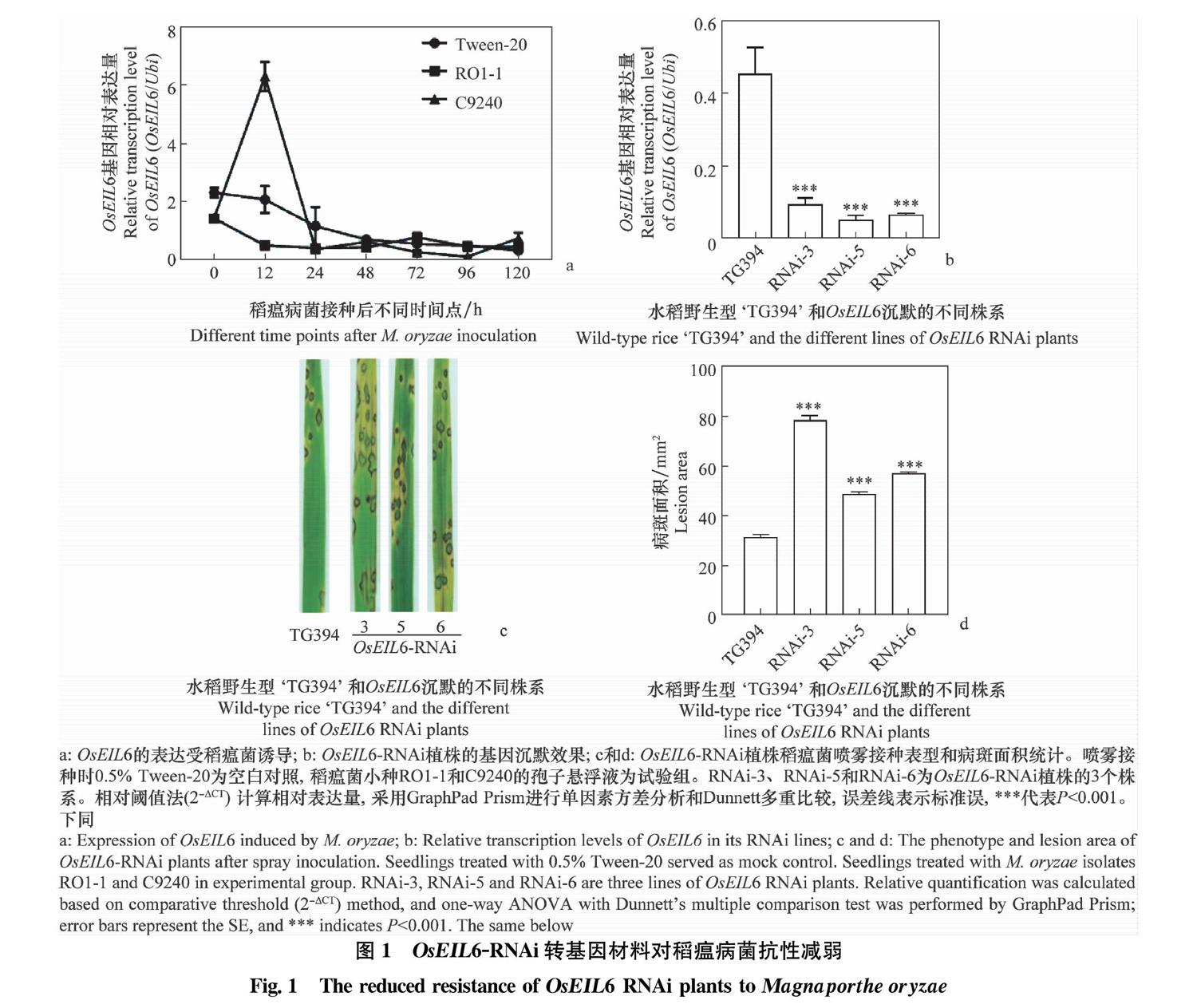
以‘日本晴為材料,采用噴霧法分別接種稻瘟病菌親和小種RO1-1和非親和小種C9240,并以清水處理為對照,檢測OsEIL6在接種0、12、24、48、72、96 h和120 h的表達情況。如圖1a所示,在非親和小種C9240接種12 h時,OsEIL6表達量顯著升高,上調約6倍;而在親和小種RO1-1接種時,OsEIL6的表達量無明顯變化。由此可見,水稻基因OsEIL6在稻瘟病菌侵染時被誘導表達。
2.2 OsEIL6-RNAi轉基因材料基因沉默效果鑒定
通過實時熒光定量RT-PCR的方法對獲得的OsEIL6-RNAi材料的基因沉默效果進行鑒定。如圖1b所示。與野生型‘TG394水稻相比,OsEIL6在RNAi轉基因材料3、5、6號株系中的表達極顯著下降,表明OsEIL6在其RNAi植株中的表達受到抑制。在繼代培養的T2代獲得了OsEIL6-RNAi轉基因純合株系,為后續的抗病性表型檢測提供了可靠材料。
2.3 OsEIL6在水稻抗稻瘟病反應中的作用
OsEIL6-RNAi轉基因材料對稻瘟菌抗性的鑒定結果如圖1c,d所示。接種稻瘟菌7 d后,與野生型‘TG394水稻相比,OsEIL6-RNAi純合株系的葉片在病斑數量和病斑面積上都有明顯的差異。OsEIL6-RNAi轉基因材料的病斑面積明顯大于‘TG394,其中發病最嚴重的株系OsEIL6-RNAi-3的葉片病斑面積可達‘TG394病斑面積的2.5倍(圖1d)。以上結果表明OsEIL6在水稻抗稻瘟病侵染過程中起著重要作用。
2.4 OsEIL6定位于水稻細胞質
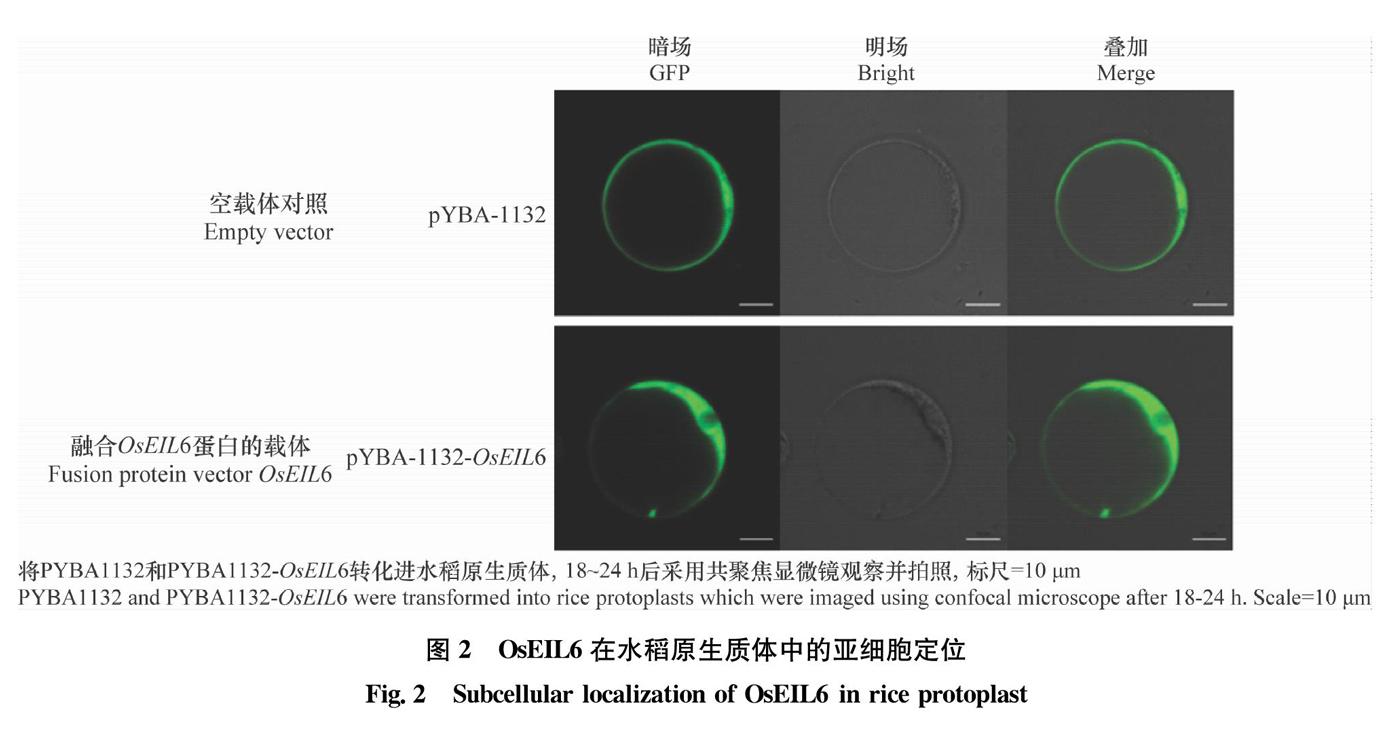
為了明確OsEIL6的功能,構建OsEIL6融合GFP標簽的表達載體pYBA1132-OsEIL6,轉化水稻原生質體后進行激光共聚焦顯微觀察。結果顯示空白載體pYBA1132的綠色熒光正常表達,并分布于全細胞(圖2)。經pYBA1132-OsEIL6侵染后的水稻原生質體,熒光特異性地出現在細胞質部位,而細胞核處未觀察到熒光信號(圖2)。該結果表明OsEIL6定位于水稻原生質體的細胞質。
2.5 OsEIL6正向調控乙烯信號途徑基因的表達
為了揭示水稻中OsEIL6可能參與的抗病信號途徑,本研究檢測了乙烯合成途徑標記基因OsACO1(1-氨基環丙烷-1-羧酸氧化酶)和乙烯信號傳導途徑標記基因OsERF063、OsERF073在OsEIL6沉默材料中的轉錄水平。結果如圖3所示,與‘TG394相比,OsEIL6基因沉默能不同程度的降低以上3個基因的表達量。其中OsACO1的表達量下降了50%~80%(圖3a),OsERF063和OsERF073的表達量也均顯著下調(圖3b,c),其中OsERF063的表達量降低了75%~80%。以上結果表明,OsEIL6正向調控乙烯信號途徑標記基因的表達。
2.6 OsEIL6正向調控水稻活性氧(ROS)合成途徑基因的表達
進一步檢測OsEIL6沉默材料中ROS合成途徑NADPH氧化酶基因OsrbohA和OsrbohB的轉錄水平,結果發現在OsEIL6-RNAi材料中,OsrbohA和OsrbohB的表達量均有不同程度的下降,雖然OsrbohB表達量的下降程度低于OsrbohA,但也達到了顯著差異水平(圖4)。上述結果表明,OsEIL6的沉默抑制活性氧合成途徑關鍵酶基因表達,進而降低水稻對稻瘟病的抗性。
2.7 OsEIL6的互作蛋白研究
本研究采用LC-MS的方法篩選OsEIL6在水稻原生質體中的互作蛋白。質譜分析結果見表2,共獲得10個可能與OsEIL6互作的候選蛋白。通過基因序列信息和功能分析,發現14-3-3蛋白和OsWRKY29都是和植物抗病性直接相關的蛋白。另外,兩個受體類蛋白質激酶(LOC_Os05g07740.1和LOC_Os05g44930.1)以及一個絲氨酸蘇氨酸蛋白質激酶(LOC_Os01g66860.4)很可能會通過磷酸化修飾調控OsEIL6的功能。而F-box類泛素連接酶(LOC_Os12g27750.1)則可通過26S蛋白酶體降解途徑調節OsEIL6的蛋白水平。后續將會進一步驗證候選蛋白與OsEIL6之間的相互作用關系,以及研究它們參與水稻抗稻瘟病反應的功能。
3 討論
在水稻6個EIN3/EIL家族基因中[12-13],OsEIL1和OsEIL2 與擬南芥EIN3的EIL1和EIL2亞族相似性最高,OsEIL3和OsEIL4與SLIM1/EIL3關系密切,可能有相同的功能,而OsEIL5和OsEIL6與報道的EIL親緣關系最遠[12]。本研究發現,OsEIL6沉默材料中乙烯合成途徑基因OsACO1以及乙烯信號傳導途徑基因OsERF063和OsERF073的表達量均顯著下降,且對稻瘟病抗性減弱,表明OsEIL6參與了乙烯信號途徑并正調控水稻抗稻瘟病。這一研究結果和已報道的OsEIL1介導的水稻抗稻瘟病的功能相一致[11, 17]。Yang等報道OsEIL1超表達植株對稻瘟病抗性增強,Helliwell等發現OsEIL1-RNAi植株對稻瘟病的抗性減弱,表明OsEIL1正調控水稻抗稻瘟病。此外,Yang等還發現OsEIL1作為轉錄因子可直接靶標NADPH氧化酶基因OsrbohA和OsrbohB的啟動子區域并激活其轉錄,從而調控ROS產生所參與的水稻對稻瘟病的抗性[18]。本研究進一步檢測了OsEIL6沉默材料中ROS合成途徑標記基因OsrbohA和OsrbohB的表達情況,發現其表達均有不同程度的下降。因此OsEIL6可能通過影響ROS的合成,進而參與水稻的抗病反應過程。
一般來說,SA信號傳導途徑能夠幫助植株有效抵抗活體營養型病菌,而植物抵抗死體營養型病菌則主要依賴于ET和JA信號途徑[19]。激素之間協同或拮抗的作用,使植物體內的信號傳導途徑更加豐富,所以當植物受到病原物侵染時可以快速精準地調控體內的防衛反應。在擬南芥中,研究發現EIN3和EIL1可直接作用于SA合成基因SID2(salicylic acid induction deficient2)的啟動子區,負調控SID2的表達和SA的合成,從而抑制植物對活體營養型病原菌的免疫反應[9]。而JA信號途徑的轉錄激活子MYC2和EIN3可相互結合,從而影響了各自的轉錄活性,導致JA和ET信號途徑間的相互拮抗作用[20]。因此,擬南芥中EIN3/EIL是控制ET和其他兩種激素JA和SA信號傳導途徑交叉的節點,并能夠調節不同激素信號途徑對不同類型病原菌的侵染做出反應。因此,研究水稻中OsEILs成員是否同樣處于調節3種激素信號傳導途徑的關鍵節點位置,以及其突變體材料對不同類型病原菌如死體營養型紋枯菌和活體營養型白葉枯菌的抗感反應,將對制定水稻的抗病育種策略具有重要意義。
水稻原生質體亞細胞定位分析發現,OsEIL6并未定位于水稻細胞核,而是定位于細胞質。這一研究結果不同于EIN3家族OsEIL1和OsEIL2的細胞核定位結果[14-15],因此,OsEIL6是否作為轉錄因子執行功能還需要后續進一步研究。蛋白質之間的相互作用是細胞生化反應網絡的重要組成部分,對調控細胞及其信號具有重要意義。本研究利用LC-MS的方法尋找OsEIL6的互作蛋白,篩選到10個可能與水稻抗病相關的互作蛋白。后續將進一步研究這些互作蛋白在水稻抗病過程中的作用,進一步解析OsEIL6所介導水稻抗病分子機制。
綜上,本研究發現OsEIL6定位于水稻細胞質,且正調控水稻對稻瘟菌的抗性反應。該結果對于進一步研究OsEIL6的生物學特性和功能,明確其在激素信號途徑中的地位以及OsEIL6所調控的水稻抗病信號分子網絡具有重要作用,并為水稻稻瘟病防控提供新思路。
參考文獻
[1] LIU Wende, LIU Jinling, TRIPLETT L, et al. Novel insights into rice innate immunity against bacterial and fungal pathogens [J]. Annual Review of Phytopathology, 2014, 52(1):213-241.
[2] TALBOT N J. On the trail of a cereal killer: Exploring the biology of Magnaporthe grisea [J]. Annual Review of Microbiology, 2003, 57(1):177-202.
[3] DEAN R A, TALBOT N J, EBBOLE D J, et al. A P-type ATPase required for rice blast disease and induction of host resistance [J]. Nature, 2006, 440 (7083): 535-539.
[4] GRANT M R, JONES J D G. Hormone (dis)harmony moulds plant health and disease [J]. Science, 2009, 324(5928): 750-752.
[5] JU Chuanli, CHANG Caren. Mechanistic insights in ethylene perception and signal transduction [J]. Plant Physiology, 2015, 169(1): 85-95.
[6] KLEE H J. Ethylene signal transduction moving beyond Arabidopsis [J]. Plant Physiology, 2004, 135(2):660-667.
[7] YANG Chao, LU Xiang, MA Biao, et al. Ethylene signaling in rice and Arabidopsis: conserved and diverged aspects [J]. Molecular Plant, 2015, 8 (4): 495-505.
[8] ZHAO He, DUAN Kaixuan, MA Biao, et al. Histidine kinase MHZ1/OsHK1 interacts with ethylene receptors to regulate root growth in rice [J/OL]. Nature Communications, 2020, 11(1):518. DOI: 10.1038/s41467-020-14313-0.
[9] CHEN Huamin, LI Xue, CHINTAMANANI S, et al. ETHYLENE INSENSITIVE3 and ETHYLENE INSENSITIVE3-LIKE1 repress SALICYLIC ACID INDUCTION DEFICIENT2 expression to negatively regulate plant innate immunity in Arabidopsis [J]. The Plant Cell, 2009, 21(8): 2527-2540.
[10]ZHU Ziqiang, AN Fengying, FENG Ying, et al. Derepression of ethylene-stabilized transcription factors (EIN3/EIL1) mediates jasmonate and ethylene signaling synergy in Arabidopsis [J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(30):12539-12544.
[11]YANG Chao, LI Wen, CAO Jidong, et al. Activation of ethylene signaling pathways enhances disease resistance by regulating ROS and phytoalexin production in rice [J]. The Plant Journal, 2017, 89(2):338-353.
[12]MAO Chuanzao, WANG Shaomin, JIA Qiaojun, et al. OsEIL1, a rice homolog of the Arabidopsis EIN3 regulates the ethylene response as a positive component [J]. Plant Molecular Biology, 2006, 61(1/2): 141-152.
[13]HIRAGA S, SASAKI K, HIBI T, et al. Involvement of two rice ETHYLENE INSENSITIVE3-LIKE genes in wound signaling [J]. Molecular Genetics & Genomics, 2009, 282(5): 517-529.
[14]石彥龍,徐國娟, 王旭麗,等. 轉錄因子OsEIL2正調控水稻對紋枯病的抗性[J].植物保護, 2017, 43(1):46-53.
[15]PARK C H, CHEN Songbiao, SHIRSEKAR G, et al. The Magnaporthe oryzae effector AvrPiz-t targets the RING E3 ubiquitin ligase APIP6 to suppress pathogen-associated molecular pattern-triggered immunity in rice [J]. The Plant Cell, 2012, 24(11):4748-4762.
[16]ZHANG Yang, SU Jianbin, DUAN Shan, et al. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes [J/OL]. Plant Methods, 2011, 7(1):30. DOI: 10.1186/1746-4811-7-30.
[17]HELLIWELL E E, WANG Qin, YANG Yinong. Ethylene biosynthesis and signaling is required for rice immune response and basal resistance against Magnaporthe oryzae infection [J]. Molecular Plant-Microbe Interactions, 2016, 29(11): 831-843.
[18]YANG Chao, MA Biao, HE Sijie, et al. MAOHUZI6/ETHYLENE INSENSITIVE3-LIKE1 and ETHYLENE INSENSITIVE3-LIKE2 regulate ethylene response of roots and coleoptiles and negatively affect salt tolerance in rice [J]. Plant Physiology, 2015, 169(1):148-165.
[19]BARI R, JONES J D. Role of plant hormones in plant defence responses [J]. Plant Molecular Biology, 2009, 69(4): 473-488.
[20]SONG Susheng, HUANG Huang, GAO Hua, et al. Interaction between MYC2 and ETHYLENE INSENSITIVE3 modulates antagonism between jasmonate and ethylene signaling in Arabidopsis [J]. The Plant Cell, 2014, 26(1): 263-279.
(責任編輯:張文蔚)
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
