白化茶樹L-茶氨酸累積機理的研究進展
2020-08-28 07:23:22疏再發王琳琳婁艷華吉慶勇邵靜娜劉瑜何衛中
食品研究與開發 2020年17期
關鍵詞:研究
疏再發,王琳琳,婁艷華,吉慶勇,邵靜娜,劉瑜,何衛中
(麗水市農業科學研究院,浙江麗水323000)
茶樹是我國重要的經濟作物,茶葉產業的健康發展關系到茶農的收入增長。茶葉產業持續、健康發展需要優異的種質支撐,特色茶葉產業化基本上以特異優異的茶樹品種引領。近些年來,具有珍貴白芽或黃芽的白化茶樹品種,在我國被廣泛種植,引領著茶樹資源的選育方向。例如浙北發現的白葉一號,成為我國綠茶新的亮點,促進了浙江省乃至全國茶葉產業的發展。白化茶因其極高的茶葉品質受到越來越多的關注,而茶葉品質的好壞受到眾多化合物綜合影響,尤其是茶葉中氨基酸,特別是茶氨酸、谷氨酸、精氨酸等茶葉品質成分,對茶葉品質起決定作用[1]。
作為茶葉中最重要的氨基酸種類—茶氨酸,在茶葉的游離氨基酸中含量最高,占總游離氨基酸的30%~60%,占茶葉干物質總量的1%~5%。茶氨酸,又稱N-乙基-γ-L-谷氨酰胺[2],是茶樹中一種獨特的非蛋白氨基酸。茶樹中天然存在的茶氨酸都屬于L-茶氨酸,L-茶氨酸是茶樹體內最重要的次級代謝產物之一,是茶葉鮮爽味的主要來源,與綠茶品質呈正相關[3]。而紅茶制作過程中游離氨基酸與糖類發生反應生成茶氨酸-糖普化合物促進紅茶茶湯紅艷和葉底紅亮[4]。茶氨酸自被發現以來,其在食品科學和保健功能等方面研究廣泛。研究表明,茶氨酸可以通過抑制咖啡因的負作用,在一定程度上具有降低血壓和調節情緒等功能[5];茶氨酸可以通過激活相關的中樞神經遞質來改善記憶和學習能力[6];茶氨酸還具有良好的抗疾病作用,包括增強抗腫瘤活性、預防血管疾病和神經保護等功能[7];茶氨酸還對人體具有鎮靜、安神等多種生理保健功能[8]。最新研究還表明L-茶氨酸能夠促進人體細胞增殖,調節葡萄糖代謝[9]。目前茶氨酸在使用過程中沒有限制用量規定,使得茶氨酸在食品和醫藥領域得到了廣泛的應用[10]。茶氨酸具有多種生理功能,而白化茶樹因其高茶氨酸備受關注,因此研究白化茶樹L-茶氨酸的累積機理具有極其重要的意義。
1 白化茶樹L-茶氨酸的差異累積
大量研究表明,大部分白化茶樹擁有更高的游離氨基酸,如白葉1號的游離氨基酸高達8.67%[2];四明雪芽的游離氨基酸含量達到9.55%[11];中黃1號的氨基酸達到8.40%[12];景白1號的游離氨基酸含量在6.90%~7.50%[13];中白4號的游離氨基酸含量在7.04%~10.54%[14]。而我國大面積種植的綠葉茶樹品種福鼎大白和龍井43的游離氨基酸含量僅為2.97%和4.99%[2]。
白化茶樹品種具有珍貴的白芽或黃芽,現已在我國被廣泛種植,因其游離氨基酸和茶氨酸的高含量累積,白化茶樹能夠生產出具有口感鮮爽、香氣濃郁等特點的高品質綠茶。Du等利用茶樹天然白化突變體選育出的小雪芽和白葉1號與綠茶生產常用品種浙農113進行比較,結果表明,3個品種游離氨基酸組分中含量最高的是茶氨酸,而兩種白化品種的總氨基酸和茶氨酸含量均顯著高于浙農113[15]。Li等對小雪芽、白葉1號和浙農113生理生化指標進行檢測,得到了相似的結論,白化茶葉綠體結構異常,總兒茶素、咖啡堿水平較低,總氨基酸特別是茶氨酸含量較高[16]。Feng等為確定白化茶品質的關鍵化學成分的變化,對4個白化茶品種和1個綠茶品種的代謝產物進行了分析。與綠茶品種相比,白化茶品種葉綠素含量(P<0.01)、總類胡蘿卜素(P<0.05)、咖啡因(P<0.01)和總兒茶素(P<0.05)顯著降低,而葉綠素a/b和游離氨基酸的含量包括茶氨酸顯著增加。多元分析表明,高水平的氨基酸,低水平的葉綠素、兒茶素和咖啡因,有助于白化茶增加鮮味而減少苦澀味[17]。Li等研究表明,不管是相對綠葉品種或紫色變異品種,白化變異品種對茶氨酸的代謝活性較高,茶氨酸在根部的含量明顯高于其他變異,在葉片中的含量明顯高于紫色變異[18]。Wang等研究白化茶樹中黃2號發現,其茶氨酸和游離氨基酸的含量明顯高于龍井43,而類胡蘿卜素、兒茶素和花青素的含量低于龍井43。微陣列分析表明,與龍井43相比,中黃2號幼苗中與氨基酸代謝、光合作用和色素代謝相關的259個基因表達都發生了顯著變化[19]。
不僅是白化茶樹品種具有高茶氨酸的特點,茶園里發現的帶有白化病的群體種茶分枝,與對照綠色葉片相比,這一特殊分枝上的白化葉片也積累了相當高的氨基酸含量,尤其是茶氨酸含量高達4.5%,而多酚含量較低,葉綠素含量也極低[20]。而Cheng等在廣東英德發現的茶樹黃色的枝條突變體,對比正常葉片(綠色),黃葉的L-茶氨酸含量明顯高于綠葉(P<0.01)。通過添加補充[2H5]-L-茶氨酸,抑制黃葉中L-茶氨酸轉化為乙胺和L-谷氨酸的酶活性,從而使其L-茶氨酸分解代謝顯著低于綠葉(P<0.05)[21]。
2 茶樹L-茶氨酸的分布及含量
Deng等研究了27種(或變種)山茶科植物葉片中茶氨酸的含量。茶氨酸存在于21個品種或變種中,但其含量(鮮重<0.2 mmol/g)遠低于茶樹品種中檢測到的含量[22]。
L-茶氨酸作為茶樹中重要且含量豐富的游離氨基酸,幾乎可以在所有組織中檢測到[23]。然而,L-茶氨酸的含量在不同茶樹品種和茶樹不同組織之間有很大的差異。一般來說,幼葉茶氨酸含量最高,其次是根、老葉和莖[4]。一年內,茶氨酸含量春季最高,隨后逐漸下降,秋季花芽完全開放成花時葉片中茶氨酸減少[24],根中茶氨酸一定程度地上升,在冬季(休眠期),茶樹中的茶氨酸積累并儲存在根部,到第二年的3月和4月(發芽期),茶氨酸再轉移到芽上,到8月時,根中的茶氨酸含量逐漸降低[4]。利用[14C]乙胺的放射性,測定了兩種茶樹幼苗中茶氨酸的生物合成活性,發現根的茶氨酸合成活性高于葉[22],而春季中茶氨酸、谷氨酰胺、丙氨酸等主要氨基酸通過木質部汁液運輸到茶樹各地上器官中[25]。研究發現,2年生茶苗培養27 d到205 d,茶氨酸分布于包括根在內的所有器官中,其中鮮葉中茶氨酸的濃度為 0.1 mg·N/g ~0.5 mg·N/g,根中茶氨酸含量為 0.5 mg·N/g~1.1 mg·N/g[26]。Liu 等開展對2個不同白化茶樹品種和1個綠葉茶樹品種的研究,檢測茶苗中一芽一葉、二葉、三葉、老葉、莖和根6個不同茶樹組織的茶氨酸含量,發現白化茶樹品種葉片和根的茶氨酸含量均高于綠葉茶樹品種,而茶樹不同組織內茶氨酸含量分布規律均為一芽一葉>根>二葉>三葉>老葉>莖,其中各茶樹品種一芽一葉茶氨酸最高為37.7 mg/gDW,而莖內茶氨酸含量最低為2.7 mg/gDW[27]。
3 茶樹L-茶氨酸合成代謝途徑關鍵酶及基因的研究
3.1 茶樹L-茶氨酸合成代謝途徑關鍵酶的研究
對于茶樹體內茶氨酸合成代謝途徑的研究,早在20世紀60至80年代,日本學者竹尾忠一通過同位素14C示蹤法對茶苗進行試驗推定茶氨酸合成的前體是谷氨酸和乙胺[28]。而后研究人員一直在研究完整的茶氨酸合成代謝途徑,目前茶氨酸合成代謝途徑如圖1所示。茶氨酸合成代謝途徑是茶樹氮代謝的重要組成部分,關鍵酶包括谷氨酸脫氫酶(glutamatedehydrogenase,GDH)、谷氨酸合成酶(glutamate synthetase,GOGAT)、丙氨酸脫羧酶(alanine decarboxylase,AlaDC)、茶氨酸合成酶(theanine synthetase,TS)、茶氨酸水解酶(theanine hydrolase,ThYD)、谷氨酰胺合成酶(glutamine synthetase,GS)及精氨酸脫羧酶(arginine decarboxylase,ADC)等[29]。其中茶氨酸合成酶、茶氨酸水解酶是茶氨酸代謝特異性的酶[16-18]。由于丙氨酸在丙氨酸脫羧酶催化下形成乙胺的反應只能在根部進行,因此盡管茶氨酸合成酶在茶樹各個器官中都存在,但茶氨酸僅在茶樹的根部合成[30]。
GDH可將植物中的碳氮代謝連接起來,因為它既能將氨同化為谷氨酸,也可將谷氨酸脫氨形成氨和α-酮戊二酸,在NH4+的合成及谷氨酸的代謝中具有十分獨特的作用[29]。GOGAT將谷氨酰胺和α-酮戊二酸轉變為2分子谷氨酸,其中一分子谷氨酸可作為GS的底物,而另一分子谷氨酸可用于合成蛋白質、核酸等含氮化合物[31]。AlaDC是一種活性蛋白,是茶氨酸合成鏈中的重要酶類,催化丙氨酸脫羧得到的茶氨酸合成前體乙胺[32]。TS是一種L-谷氨酸-乙胺連接酶,是茶氨酸合成的關鍵酶,對乙胺具有高親和力,在腺苷三磷酸(adenosine triphosphate,ATP)、K+、Mg2+的催化下,可將乙胺和L-谷氨酸合成L-茶氨酸[33]。茶氨酸水解酶屬水解酶家族,作用于碳氮鍵而不是肽鍵,在茶樹體內,茶氨酸的水解主要是由茶氨酸水解酶和谷氨酰胺酶催化,被分解形成谷氨酸和乙胺[34]。GS是無機氮到有機氮同化利用的關鍵催化酶,有兩種同工酶,即分別定位于細胞質和葉綠體的GS1和GS2,它們分別執行不同的生理功能。位于葉綠體中的GS2的主要功能是把葉綠體和光呼吸再合成的NH4+合成為谷氨酰胺,而分布在根部的GS1則主要參與根部氮的合成,在ATP存在的情況下,催化谷氨酸和氨合成谷氨酰胺[35]。ADC在植物的生長、花芽分化、逆境脅迫響應等多種生理活動中發揮著關鍵作用,主要是催化精氨酸形成鯡精胺[36]。
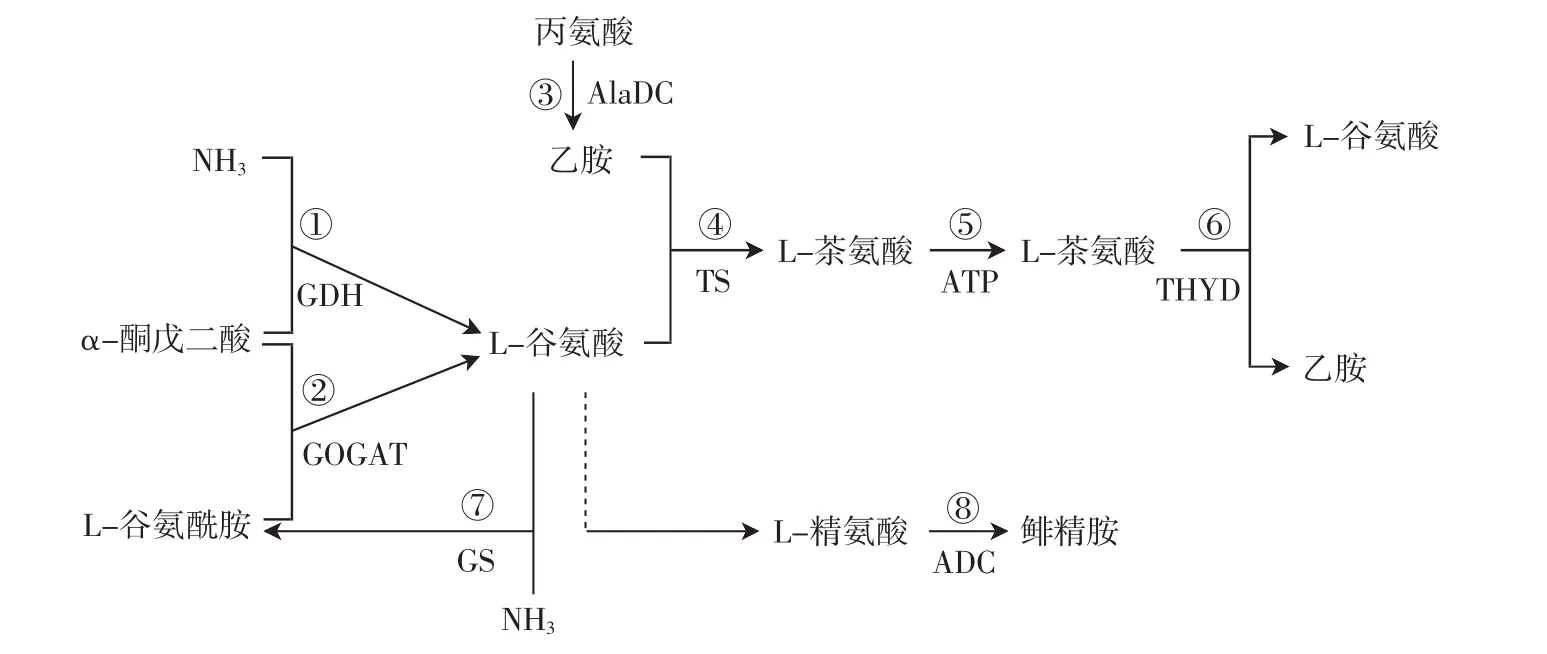
圖1 L-茶氨酸合成代謝途徑關鍵酶Fig.1 Key enzyme of L-theanine anabolism pathway
3.2 茶樹L-茶氨酸合成代謝途徑相關基因的研究
參與茶氨酸生物合成途徑的關鍵基因包括GDH、GOGAT、AlaDC、TS、GS 和 ADC,分別代表編碼谷氨酸脫氫酶、谷氨酸合成酶、丙氨酸脫羧酶、茶氨酸合成酶、谷氨酰胺合成酶和精氨酸脫羧酶的基因。已克隆的L-茶氨酸合成代謝途徑相關基因序列信息如表1所示。陳琪等研究表明TS在茶樹不同器官中均有表達,其在根系、芽、花中表達相對較高;而GOGAT和GDH在葉片和根系表達,其中GOGAT葉片表達量高于根系,而GDH剛好相反,這是因為兩者酶功能間的差異[37]。Wei等在添加乙胺前體培養茶樹的基礎上研究茶氨酸生物合成和關鍵基因表達的途徑,結果顯示CsTS1和GS2-1.1的表達模式與組織中茶氨酸含量顯著相關,根中CsTS1表達在處理12 d后上升到最高水平,同時茶氨酸積累達到高峰。而 GS、TS、GOGAT、GDH 和ADC基因的表達顯著增加,谷氨酸、茶氨酸和精氨酸含量升高,結果均表明這些基因參與茶氨酸積累,且是茶樹體內氮同化、儲存和循環的一部分[38]。

表1 茶氨酸合成途徑相關酶基因Table 1 Related gene in theanine synthesis pathway
茶氨酸的合成代謝途徑,包括合成、運輸和水解,許多關鍵酶參與茶氨酸的合成代謝途徑,而這些合成代謝關鍵酶基因表達水平在很大程度上影響茶氨酸在茶樹中的含量、分布和代謝規律。在茶氨酸生物合成中,許多結構基因編碼酶及酶基因共同作用,從而直接表達催化導致茶氨酸形成此一系列的反應步驟。
關于白化茶樹茶氨酸的合成代謝關鍵酶基因的研究,Liu等研究兩個白化茶樹品種和綠葉品種的不同組織中茶氨酸合成代謝關鍵基因的差異,結果表明,隨著葉片成熟度的增加,CsTS1和GOGAT的轉錄豐度增加,而CsTS2、CsGS1和CsGDH2的轉錄豐度降低;CsGS2、GOGAT的轉錄水平在芽期和第1葉、第2葉、第3葉時逐漸升高,但老葉較低;ADC在3葉的表達量最高,其次為芽、1葉、2葉、老葉;在所選基因中,莖和根的轉錄豐度較低;CsTS1、CsTS2、CsGS1、CsGS2在根中轉錄豐度最低;CsGS1、CsGS2的轉錄水平接近于零。CsTS2、CsGDH1、ADC、CsTS2、CsGDH1 的轉錄水平在白化茶樹組織中最高[27]。Lu等在研究茶園中白化變異葉片發現,白葉葉片中游離氨基酸特別是谷氨酸和茶氨酸的含量高于綠葉葉片,而白化葉中茶氨酸合成酶(TS)的基因表達量并沒有明顯增加,谷氨酰胺合成酶(GS)的表達明顯上調[20]。Cheng等比較了白化變異葉片和正常的綠葉中這3個TS基因的表達水平,白化誘導的黃葉中CsTS1和CsTS2的表達量在春季和冬季均明顯低于正常綠葉,而CsTS1轉錄量在兩季中均無顯著差異[21]。這些結果均說明,茶園中發現白化變異的黃葉的茶氨累積與L-茶氨酸生物合成的被激活無關。
4 茶樹L-茶氨酸的合成代謝的影響因素
4.1 施肥管理對茶氨酸合成代謝的影響
4.1.1 N、P、K對茶氨酸合成代謝的影響
N是茶樹生長最重要的肥源之一,N肥供應量與茶樹體內組織含氮量息息相關,而茶樹組織含氮量與茶樹體內茶氨酸含量存在明顯的相關性。大量研究表明,茶樹偏銨態氮吸收,銨態氮濃度與茶葉茶氨酸含量存在正相關,高銨態氮可以激活茶氨酸合成的GDH途徑,形成谷氨酰胺、茶氨酸等物質[40]。Yang通過15N標記和動力學試驗表明,與15NO3-相比,添加15NH4+的茶樹積累了更多的同化氮,促進谷氨酰胺、茶氨酸等合成[41]。P占茶樹全株干重含量的0.3%~0.5%,與茶樹體內氨基酸含量密切相關[42]。林鄭和通過不同P濃度處理的研究表明,隨著P濃度的升高,茶氨酸含量呈現不斷升高的趨勢,相對 P 濃度為 200 μmol/L~400 μmol/L,不供 P處理下茶氨酸含量顯著降低,而P濃度為1 000 μmol/L處理下茶氨酸含量顯著增加[43]。K占茶樹全株干重含量的0.5%~1.0%,也與茶樹體內氨基酸含量密切相關,土壤鉀元素能夠明顯提高茶葉內游離氨基酸含量[44],施鉀肥處理茶氨酸含量相對于不施鉀肥明顯升高,但鉀肥水平高低與茶氨酸含量高低不存在明顯相關[45]。
N、P、K作為植物三元素,是茶樹施肥補充的主體。楊婷婷通過土壤中添加Se和Zn與N和/或P的研究發現,單獨添加P處理一芽二葉和老葉的氨基酸提高17.3%和5.9%,單獨添加N處理一芽二葉和老葉的氨基酸提高37.3%和14.5%,添加P+N處理一芽二葉和老葉的氨基酸提高26.1%和13.2%,各處理間均差異顯著[46]。羅凡等研究表明,配施N+P+K肥,春茶氨基酸總量和茶氨酸含量在單施 N、P、K、N+P、N+K、P+K中含量最高,其含量比對照不施肥處理提高27.88%和26.60%,差異顯著。說明氮、磷、鉀配施在各組分共同作用下有利于提高春茶氨基酸總量及茶氨酸含量,從而提高春茶的品質[47]。
4.1.2 微量元素對茶氨酸合成代謝的影響
微量元素是茶樹生長發育中不可或缺的養分,影響茶樹茶氨酸合成代謝的進行。Ruan等采用營養液、土壤盆栽和田間試驗研究了鎂營養狀況對茶樹游離氨基酸積累和轉運的影響,結果表明在不影響葉片總氮、無機氮吸收量和谷氨酰胺合成酶活性的前提下,營養液中Mg2+的充足供應可以提高生物量和游離氨基酸的濃度,特別是芽和根中的茶氨酸[48]。而茶樹是聚錳植物,錳有利于茶樹進行光合作用,有利于促進茶樹氮代謝,有利于茶樹根系進行硝態氮還原作用,進而合成氨基酸。茶樹施用錳后,春茶茶葉中的氨基酸增加34.75%[49]。而鉬是硝酸還原酶的重要組成成分,茶樹缺鉬情況下氨基酸合成受阻,茶園施用鉬肥能夠顯著增加茶氨酸含量(比對照增加49%)[50]。研究發現鋅也與硝酸還原酶活性相關,施鋅肥能夠明顯提高茶樹體內氮和氨基酸含量[51]。趙萌用云抗10號茶樹品種作為研究材料,發現茶氨酸含量隨著土壤施鋅量的增加而不斷增加,在施鋅量為24 mg/kg時,茶氨酸含量為1.7%,較不施肥提高57.4%;同時茶氨酸含量還隨著葉面噴施銅肥的增加而增加,而在施銅量為300 mg/kg時茶氨酸含量為1.68%,較對照增幅達到最大的44.44%[52]。王麗霞利用平陽特早(高氟品種)和福鼎大白(低氟品種)作為研究材料發現,氟處理增加平陽特早的茶氨酸含量而降低福鼎大白的茶氨酸,這可能與茶樹體內的揮發物質有關[53]。Zhu等通過F處理龍井長葉,研究發現多酚含量、游離氨基酸、總兒茶素明顯下降,但是茶氨酸含量與游離氨基酸呈相反的趨勢,在2 d后達到高峰[54]。
4.2 環境因素對茶氨酸合成代謝的影響
4.2.1 溫濕度對茶氨酸合成代謝的影響
溫濕度影響茶樹茶氨酸的合成代謝。研究發現不同溫度下茶苗新梢內游離氨基酸隨溫度的升高而減少,且差異均達到顯著水平(P<0.05)。低溫(5℃/15℃)條件下新梢中游離氨基酸含量高達3.67%,高于中溫(10℃/20℃)條件下的3.38%和高溫(15℃/25℃)條件下的3.09%[55]。Li等采用大棚試驗和大田試驗研究了鐵觀音茶樹對虧缺水源(deficit irrigation,DI)和嚴重虧缺水源(severe deficit irrigation,SDI)的生理反應,并以全虧(full irrigation,FI)和非虧缺(non-irrigation,NI)為對照。DI在大田茶樹葉片水勢維持在-2.0 MPa~-2.5 MPa的中度水分脅迫,而SDI田茶樹葉片水勢的水分脅迫越來越嚴重。DI對比FI對茶葉鮮重和干重的影響相似,但增加了茶氨酸含量[56]。水分脅迫是影響茶樹產量和品質最重要的環境脅迫之一。Wang等研究干旱脅迫對茶葉品質的影響,干旱脅迫顯著降低茶葉中總多酚和自由氨基酸的含量,特別是茶氨酸和一些游離氨基酸顯著降低[57]。
4.2.2 光照對茶氨酸合成代謝的影響
不同光質影響茶樹茶氨酸合成代謝,研究表明茶氨酸生物合成途徑中有較多的基因在各種單色光下差異表達,相對于在紫光或黃光條件下,更多茶氨酸生物合成相關基因在藍光條件下表達量降低,這說明茶氨酸途徑對光質量的變化敏感,且對藍光最敏感[58]。不同的光照強度也影響著茶樹茶氨酸合成代謝,張昆研究發現遮光處理鮮葉氨基酸含量顯著高于不遮光處理,增幅分別為0.29%~0.53%,且遮光率越高,氨基酸含量越高[59]。Tomohito Sano等研究不同覆蓋密度和覆蓋時間下碾茶中茶氨酸的變化,研究發現不覆蓋下茶氨酸含量不斷降低,而遮光率為75%和90%下茶氨酸含量隨著覆蓋時間的延長表現出先升高后降低的規律,其中覆蓋15 d茶氨酸含量最高[60]。等對韓國抹茶覆蓋研究中也表現出相似的變化規律[61]。遮陰可以有效地提高茶葉的品質,通過增加茶氨酸的濃度,有助于茶葉新鮮和爽口的味道。研究分析了遮陰對茶氨酸生物合成的影響,以及遮陰對茶葉中茶氨酸合成相關基因表達的影響,發現遮陰對幼葉中茶氨酸和總游離氨基酸的含量都有顯著的影響。實時熒光定量 PCR(real-time quantitative PCR,qPCR)和 Western Blot分析結果表明,GS、GDH、GOGAT、NiR、ADC 等所有與茶氨酸通路相關基因的信使RNA(messenger RNA,mRNA)表達均受遮陰顯著影響。遮陰處理的老幼葉NiR和ADC基因表達均明顯降低,說明遮陰處理可能抑制了氮代謝。因此遮陰處理可以提高茶葉中游離氨基酸特別是茶氨酸的含量,提高夏茶的茶葉品質[62]。研究還發現,在日照減少的氣候條件下種植的茶葉,L-茶氨酸和咖啡因的濃度更高,而兒茶素的含量更低[63]。
4.2.3 生態環境對茶氨酸合成代謝的影響
茶氨酸的含量會隨著不同的生態因素而變化,包括種植地點、種植海拔等。例如茶氨酸水平在斯里蘭卡的茶樹品種含量為1.38 mg/g~24.18 mg/g,而肯尼亞的茶樹品種含量為2.0mg/g~17.0mg/g[63]。Han等收集了中國東部江西省廬山5個地點(海拔212 m~1 020 m不等)的綠茶樣品。結果表明,種植海拔越高,游離氨基酸含量越高,尤其是茶氨酸含量,隨著海拔梯度的增加而增加[64]。茶樹間作為茶樹生長營造更好的生態環境,對茶葉品質有著顯著影響,Ma等通過研究不間作、低密度間作和高密度間作3種環境下茶樹內含物質的變化,發現間作增加了茶芽的長度和重量,降低了氨基酸和兒茶素的含量,但增加了茶氨酸的含量[65]。
5 展望
現階段茶葉產業依舊飛速發展,這得益于茶葉的保健功效,特別是茶葉中的茶多酚、咖啡堿和游離氨基酸等,其中茶氨酸為人體非必需氨基酸,是一種安全、無毒的非蛋白氨基酸,具有豐富的保健作用,其中包括抗疲勞作用、鎮靜作用、保護腦細胞作用、抗腫瘤作用、降血壓作用等[7]。現如今茶氨酸作為功能成分添加在食品中,且在使用過程中沒有限制用量規定,茶氨酸在疾病防御及療效上得到了廣泛深入地研究。但是目前的研究多是集中在實驗動物領域,在臨床醫學上的功能和療效缺乏必要地研究,因此未來在加強開發新的茶氨酸功能性產品的基礎上,需要深入探索茶氨酸在臨床醫學上的功能和療效。
白化茶樹是一種自然變異的茶樹種質資源,大部分白化茶樹具有更高的游離氨基酸和L-茶氨酸,因此白化茶樹選育趨向于高L-茶氨酸,研究白化茶樹L-茶氨酸的累積機理具有重要的意義。白化茶樹的白化機理分為生態敏感型和生態不敏感型兩種,生態敏感型白化茶樹品種一是溫度敏感型茶樹品種,如白葉1號、小雪芽等;二是光照敏感型茶樹品種,如金光、郁金香等[66]。未來可以通過分析白化茶樹的白化機理,進而從生態因子、生理生化、分子生物學等方面開展茶樹L-茶氨酸的合成代謝及相關調控網絡解析研究,最終研究白化茶樹白化階段的L-茶氨酸的高累積機理。
陳亮等在《農作物優異種質資源評價規范(茶樹)》中明確茶氨酸≥3%(茶樹蒸青樣)為高茶氨酸茶樹種質資源,目前高茶氨酸的茶樹種質資源數量相對較少,對于高茶氨酸的茶樹品種選育就顯得尤為迫切。對于高茶氨酸的茶樹品種選育需要常規茶樹育種技術與分子標記輔助育種相結合,一是通過調查在自然群體中發現白化變異資源開展系統選育;二是通過雜交技術利用已有的高氨基酸茶樹資源相互雜交進行雜交選育;三是通過分子標記進行高茶氨酸潛在資源的篩選進行分子標記輔助育種。分子標記輔助育種可以加快育種進程,通過數量性狀基因座(quantitative trait locus,QTL)定位技術獲得高茶氨酸QTL基因位點,基于茶樹基因組數據庫和第3代分子標記的基因關聯分析,在常規育種開始前縮小育種范圍,進一步提高育種效率,從而選育更多高L-茶氨酸的茶樹品種,最終大力促進茶葉產業發展。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
