黃金梨葉片凈光合速率與生理生態因子的關系
2020-08-29 07:32:18晁新勝齊紅燕康玉柱王傳印王防震
山東林業科技 2020年4期
晁新勝,齊紅燕,康玉柱,王傳印,王防震
(1.菏澤市行政審批踏勘評審中心,山東 菏澤274000;2.菏澤市定陶區林業局,山東 菏澤274000;3.菏澤市林業局,山東 菏澤274000;4.牡丹區林業局,山東 菏澤274000)
黃金梨(Pyrus pyrifolia ‘Hwangkeumbae’) 屬沙梨系統,其果實呈圓形、果皮為金黃色,以肉嫩多汁、味甜酥脆的極佳風味而著稱[1]。目前在我國栽培已具有一定規模。光合作用既是形成果樹產量的來源,又是提高果實品質的基礎,研究黃金梨的光合作用,對于改進栽培措施、提高產量和品質具有重要意義。迄今為止,對于黃金梨的研究主要集中在生物特性觀察、栽培管理、果實貯藏,以及施肥等對光合特性的影響等方面,并取得了大量研究成果。本研究通過園內觀測其光合特性,并分析探討其與相關環境、生理因子的關系,以期為黃金梨生產栽培提供理論依據與參考。
1 材料和方法
1.1 材料
供試品種為5年生黃金梨樹,該園地處黃河故道地區,肥力中等,株行距為3m×4m,紡錘形樹體結構。選擇長勢中等、樹冠大小基本一致的果園內樹體作為研究對象。
1.2 測定方法
利用LI-6400 便攜帶式光合測定儀,于5月初的晴朗無風天氣,測定光合指標,測定時間為每天從8:00-18:00,每1h 測定1 次。
選取3 株樹,每株選3 片枝條中部的1年生成熟葉片進行測定,被測定的葉片掛牌標記,每個葉片3 次重復,記錄每片葉測試數據。
1.3 數據處理
用Excel、SPSS、STAT 處理所得數據,以平均值進行統計和分析。
2 結果與分析
2.1 黃金梨葉片凈光合速率Pn 的日變化
由圖1可看出,黃金梨葉片Pn 的日變化呈雙峰型曲線,在8:00-11:00 之間,Pn 呈上升趨勢,且上升速度較快,11:00 左右出現第一個高峰,并為全天最高峰,峰值為9.10 μmol CO2·m-2·s-1;在11:00-13:00,Pn 持續下降,至13:00 左右時,出現低谷值,此時Pn為第一次峰值的84.40%;而后又開始緩慢上升,至14:00 左右出現第2 個高峰,峰值為8.30 μmol CO2·m-2·s-1;14:00 后,葉片凈光合速率開始出現下降趨勢,至18:00 時達到最低值。葉片凈光合速率的上午峰值為下午峰值的109.64%,一天內凈光合速率的極差達6.41μmol CO2·m-2·s-1,具有光合“午休”現象。
午間氣溫過高、水分脅迫、光合有效輻射過高等均可引起植物光合“午休”。從本試驗可看出,引起黃金梨光合“午休”現象的最可能是因光合有效輻射(PAR)和大氣溫度過高所致。由于中午光照強,為減少蒸騰作用引起的水分流失,部分葉片氣孔關閉,吸收的二氧化碳減少,胞間二氧化碳濃度降低,導致光合作用減弱;而13:00 時后,隨著光照強度的逐漸降低,大氣溫度(Ta)下降,空氣相對濕度(RH)上升,氣孔又重新打開,使光合作用又逐漸增強,14:00 時達到第2 次高峰。

圖1 葉片凈光合速率日變化Figure 1 Diurnal changes of Pn in leaves of Hwangkumbae
2.2 黃金梨田間主要環境因子、生理因子的日變化
2.2.1 環境因子日變化
從圖2看出,光合有效輻射(PAR)從8:00 開始,快速上升,且上升幅度較大,至13:00 左右達到峰值,隨后開始呈下降趨勢。大氣溫度(Ta)從8:00 開始也呈上升趨勢,但上升幅度較小,至14:00 左右達到峰值,而后開始下降。空氣相對濕度和大氣二氧化碳濃度的變化趨勢相反,呈現出先降后升的趨勢,空氣相對濕度(RH)波動幅度較小,于13:00 降低至最低值。大氣二氧化碳濃度上下波動較大,最低值與最高值相差16.99 μmol CO2·mol-1。
與凈光合速率(Pn)日變化(圖1)相比較,PAR 在8:00-11:00 和14:00-18:00 兩個時間段的變化趨勢與Pn 一致;而在11:00-14:00 之間,二者變化趨勢相反;大氣二氧化碳濃度(Ca)與凈光臺速率(Pn)在11:00-14:00 之間變化相比,變化幅度較小;比較PAR、Ta 的峰值,以及RH、Pn 的最低值,出現時間相近。觀測結果表明,黃金梨Pn 與PAR、Ta 和RH呈正相關,且光合“午休”現象的最直接原因可能是由于光照強度和大氣溫度過高導致。
2.2.2 黃金梨葉片主要生理因子的日變化
如圖3所示,氣孔導度Gs 和蒸騰速率Tr 二者的日變化與Pn 相似,曲線變化均呈雙峰型,升降變幅較大。細胞間隙CO2濃度Ci 表現為先呈緩慢下降趨勢然后逐漸上升。從圖1和圖3可以看出,在11:00 前,黃金梨葉片Ci 隨著Pn 的增強反而呈下降趨勢。在11:00-12:00,Pn、Ci、Gs 均呈下降趨勢,在此期間葉片部分氣孔受較強的PAR 誘導而關閉,進入葉片的二氧化碳變少,從而導致Ci 降低。在12:00-13:00,葉片的Pn 和Gs 持續下降,Ci 略有上升,在這段時間內Ci 不再是導致Pn 下降的直接原因,主要是由非氣孔因素控制Pn 的變化。
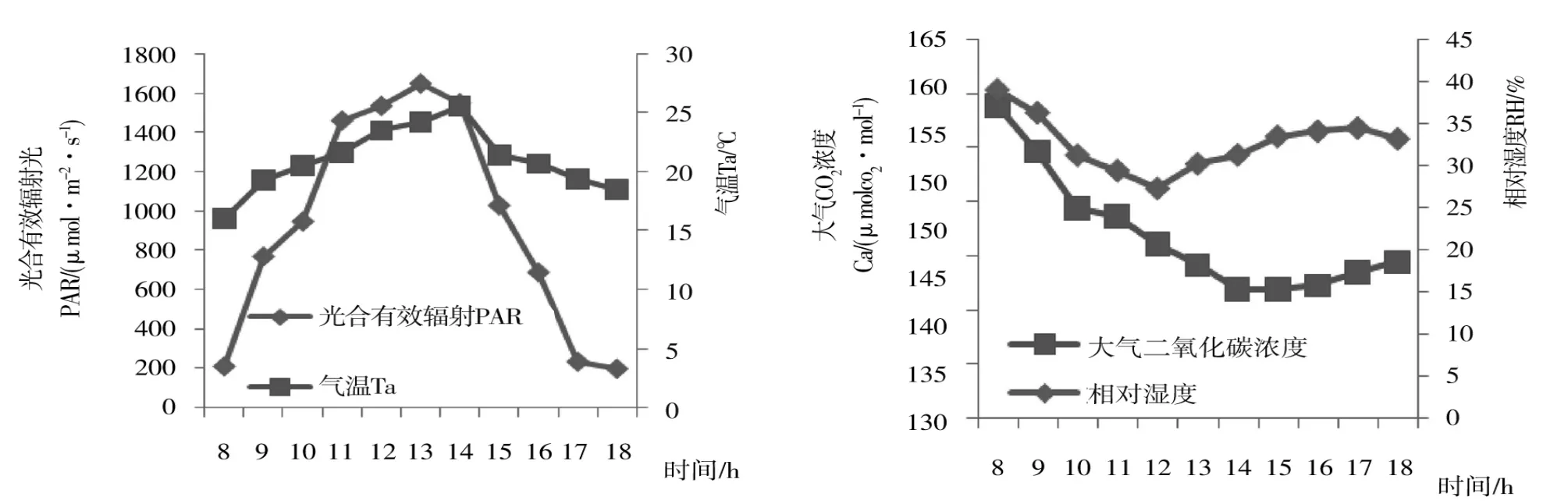
圖2 田間光合有效輻射、氣溫、相對濕度和大氣CO2 濃度的日變化Figure 2 Diurnal changes of PAR,Ta,RH and Ca under field condition
日出后隨著PAR 的增強,葉片Pn、Gs 和Tr 增大,11:00 左右葉片Gs 和Tr 分別出現第1 個高峰,峰值為0.13 mol H2O·m-2·s-1和2.48 mmol H2O·m-2·s-1,與葉片Pn 第1 個高峰出現時間相一致。隨著光照強度的增大和氣溫的升高,葉片溫度升高,氣孔部分關閉,氣孔導度減小,蒸騰速率降低,13:00 前后達到低谷,降幅為85%。13:00 后,隨著PAR 和Ta 的逐漸下降,空氣濕度升高,蒸騰速率開始開始呈現上升趨勢,14:00 左右Gs 和Tr 出現次高峰,峰值分別為0.99 mol H2O·m-2·s-1和2.68 mmol H2O·m-2·s-1。此后,由于光強和溫度的逐漸降低,Gs 和Tr 開始呈下降趨勢。這說明,Gs 和Tr 與Pn 有一定的正相關關系。

圖3 黃金梨葉片Ci、Gs 和Tr日變化
3 小結與討論
3.1 小結
黃金梨葉片凈光合速率日變化呈現雙峰曲線,兩次高峰分別出現在上午11:00 和下午14:00,具有典型的光合午休現象;黃金梨葉片凈光合速率Pn 與果園的PAR、Ta 和RH 密切相關,Pn 與Gs和Tr 有一定的正相關關系。對黃金梨葉片凈光合速率起主要作用的環境因子是光合有效輻射和相對濕度,以及氣孔導度和蒸騰速率等生理因子。
3.2 討論
光合作用的強弱對果樹的產量和品質的高低有很大影響,前人已經在梨、蘋果、核桃等果樹展開了大量的研究。果樹葉片的凈光合速率日變化曲線主要有三種類型。對于大部分果樹來說,其光合速率日變化規律呈雙峰曲線[2]。
光照是植物進行光合作用的必要條件,光合速率的高低受光直接影響。光合作用中的暗反應受相關酶所催化,而溫度是酶的活性直接影響,因此光合作用受溫度影響也很大。莫新祿[3]在砂梨上的研究表明影響光合作用的另一個是空氣相對濕度。周雷等[4]在研究春季氣象條件對溫州蜜桔光合作用的影響時指出,當相對濕度為73%時,柑橘光合作用最為適宜,低于65%或者高于81%時,濕度對光合作用起抑制作用。
氣孔是植物與外界進行氣體交換的孔道和控制蒸騰的結構,氣孔導度(Gs)的大小直接影響凈光合速率(Pn)的高低和細胞間二氧化碳濃度(Ci)的大小。馬媛等[5]在不同杏的光合特性研究中發現,Gs 極易受環境因子影響,當光強和溫度不適宜時氣孔會關閉,胞間二氧化碳濃度降低,導致中午的光合速率上升趨勢停止。
從凈光合作用曲線可發現,黃金梨凈光合速率日變化呈雙峰不對稱型曲線,并具有光合“午休”現象。這與其他學者在核果類果樹上研究結果保持一致。
通過實驗數據與圖形我們可以發現光合有效輻射和大氣相對濕度是影響黃金梨葉片Pn日變化的主要環境因子,氣孔導度和蒸騰速率是主要生理因子。這對我們在黃金梨栽培管理中果實品質的提高有很大的意義。劉遵春等[6]在研究金光杏梅葉片凈光合速率與生理生態因子的關系時指出,植物光合作用的是一個相當復雜的過程,它是由內外各種因子共同決定的。在生產中,我們要想通過改善植株的光合作用,實現促進生產的目的,必須從整體上把握,要把某個(某些)生理或生態因子放到整個植株的生態環境中去,并考慮其植株內在因素的影響,這樣才能更好地達到生產的目的。
