山東晚熟桃的SCoT遺傳多樣性分析
2020-08-31 08:44:28李淼李桂祥劉偉董曉民高曉蘭滕興榮張守民徐志芳王孝友張安寧
山東農業科學 2020年7期
關鍵詞:分析
李淼,李桂祥,劉偉,董曉民,高曉蘭,滕興榮,張守民,徐志芳,王孝友,張安寧
(1.山東省果樹研究所,山東 泰安 271000;2.沂源縣果品產銷服務中心,山東 淄博 255000;3.蒙陰縣果業發展服務中心,山東 臨沂 276500)
桃(Amygdalus persicaL.)屬薔薇科桃屬植物,落葉小喬木,原產中國。山東省是全國桃主產區,多年來栽培面積和產量穩居全國之首。桃肉質鮮美,營養豐富,深受消費者喜愛。隨著桃產業的蓬勃發展,桃品種更新替代加快,越來越多的新品種涌入市場,滿足并豐富果農的選擇。但是,桃品種缺少一套科學嚴謹的分類方法,如何快速、準確對桃品種進行有效鑒定和分類成為亟需解決的問題。
DNA分子標記技術作為鑒定作物品種的主要方法之一,在指紋圖譜構建、遺傳關系鑒定中應用較為成熟。SCoT(start codon targeted polymorphism)分子標記,即目標起始密碼子多態性標記,是一種依據植物基因中的ATG翻譯起始位點側翼序列的保守性,設計單引物并對基因組進行擴增的新型目的基因分子標記技術[1]。SCoT有別于AFLP[2]、ISSR[3]、SSR[4]、RAPD[5]等傳統意義上的隨機分子標記,以其成本低、引物通用性強、易操作和重復性好等多方面優勢在多種作物上得到廣泛應用,如獼猴桃[6]、柑橘[7]。
本研究采集山東桃產區有代表性的中晚熟桃和極晚熟桃,建立SCoT-PCR體系進行桃遺傳多樣性分析,為桃種質鑒定工作提供理論依據,并為果農選擇品種時提供參考。
1 材料與方法
1.1 材料
供試11個桃品種分別為秋風蜜、金秋紅蜜、肥城桃、八月脆、寒露蜜、青州蜜桃、沂蒙霜紅、映霜紅、中華壽桃、金牛山1號和晶白桃,詳見表1。
于2019年6月18—20日采集不同桃品種成熟葉片,每種采摘10片混合均勻,用錫箔紙包裝后放入液氮,盡快帶回實驗室于-80℃保存備用。
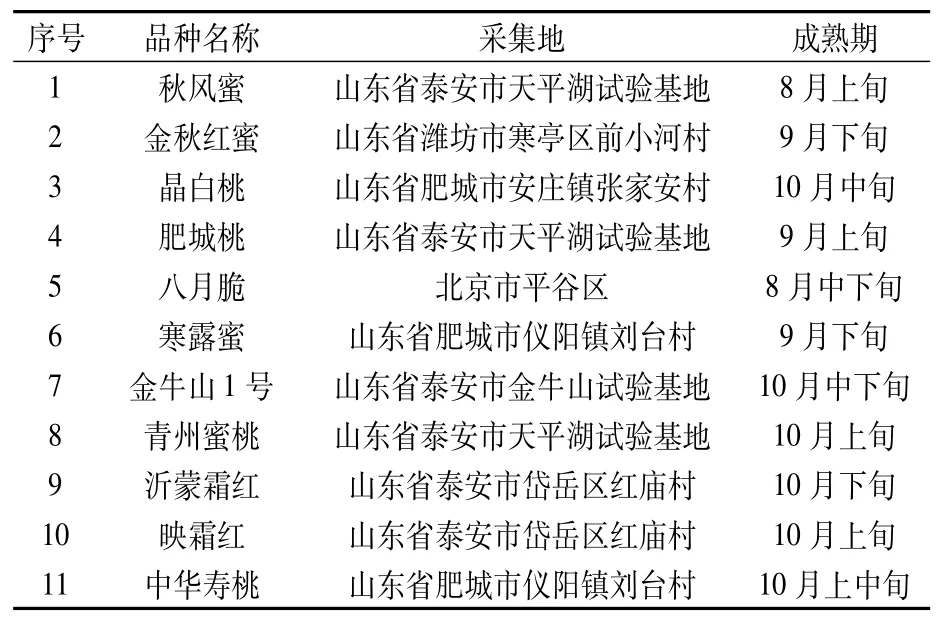
表1 試驗用桃品種詳細信息
1.2 試驗方法
1.2.1 DNA提取 采用CTAB法[8]提取桃葉DNA,并用NanoDrop One微量核酸蛋白濃度測定儀檢測其濃度和純度,最后統一稀釋至30 mg/L,置于-20℃保存備用。
1.2.2 SCoT引物及PCR擴增 本試驗所用引物均來自植物研究中已公布的SCoT引物(表2)。SCoT-PCR反應體系(20μL):2.5 mmol/L Mg2+1 μL、0.3 mmol/L dNTPs 0.4μL、30 mg/L DNA模板1 μL、1.00μmol/L引物0.5μL、0.4 U Taq DNA聚合酶0.5μL、ddH2O 16.6μL。擴增程序:94℃預變性5 min;94℃30 s,48℃30 s,72℃2 min,35個循環;72℃延伸10 min。PCR產物用1.8%瓊脂糖凝膠電泳進行檢測,并在ChamlGelTM6000型凝膠成像系統上觀察并拍照。

表2 SCoT引物序列信息
1.3 統計分析
對不同樣品同一位置電泳膠條帶有無進行賦值,有條帶記為1,無條帶記為0,形成(0,1)矩陣。采用NTSYS pc 2.1軟件計算不同品種間的遺傳相似系數,利用UPGMA法進行聚類分析,并根據遺傳相似系數進行主成分分析(PCA)。
2 結果與分析
2.1 SCoT-PCR擴增產物的多態性分析
由表3可以看出,14條引物共擴增出170條清晰、整齊的條帶,平均每條引物可擴增12.14條。其中,多態性條帶共118條,多態性比率為50.00%~93.33%,平均多態性比率為68.43%。引物SCOT8對11個桃品種的擴增結果見圖1。
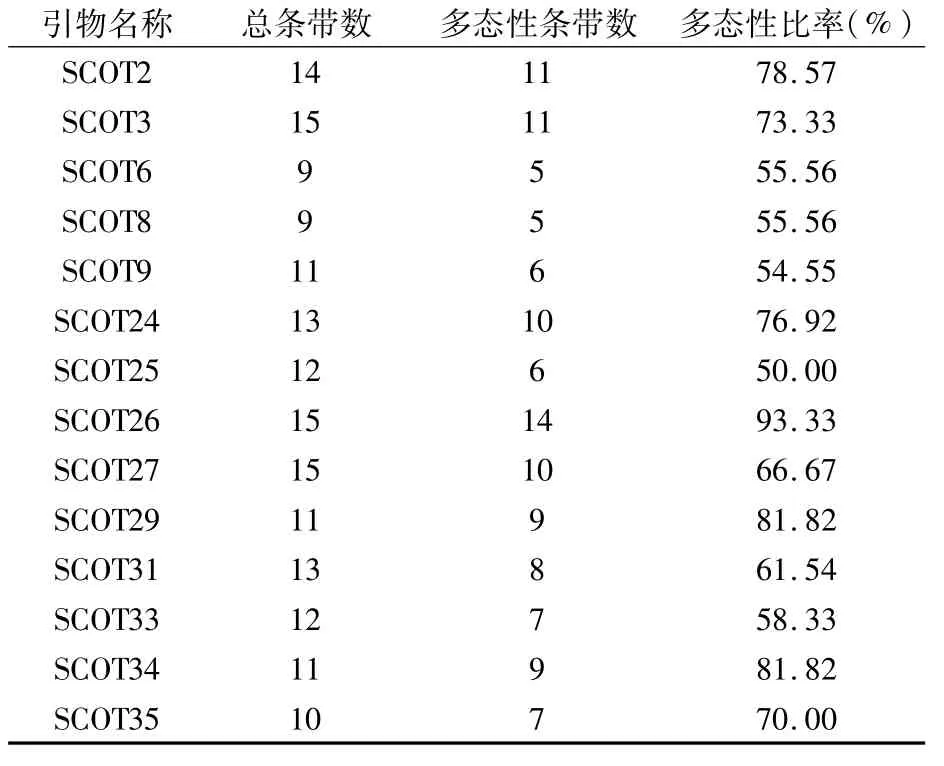
表3 SCoT-PCR擴增結果
2.2 遺傳距離及其遺傳多樣性分析
根據SCoT-PCR擴增電泳結果所轉換的(0,1)矩陣表,利用NTSYS pc 2.1軟件計算各品種間的遺傳相似系數。由表4可知,11個桃品種間的遺傳相似系數在0.37~0.89之間,平均值是0.69。其中沂蒙霜紅和映霜紅、金牛山1號和青州蜜桃、金牛山1號和映霜紅間的遺傳相似系數均較大,為0.89,表明其親緣關系較近。秋風蜜和肥城桃的遺傳相似系數最小,表明兩者親緣關系較遠,因此秋風蜜具有肥城桃的遺傳背景的可能性較小。11個桃品種間遺傳相似系數分布0.75以上的組合有21個,占總體的38.18%,遺傳相似系數小于0.75的品種占61.82%。綜上可知,11個桃品種間的遺傳相似性較低,遺傳多樣性較高。
2.3 聚類分析和主坐標分析
由圖2可以看出,在遺傳相似系數為0.72水平上,可以將11個桃品種分為3組。第Ⅰ組為秋風蜜和八月脆;第Ⅱ組包括7個品種:金秋紅蜜、寒露蜜、青州蜜桃、沂蒙霜紅、映霜紅、中華壽桃、金牛山1號;第Ⅲ組包括肥城桃和晶白桃。
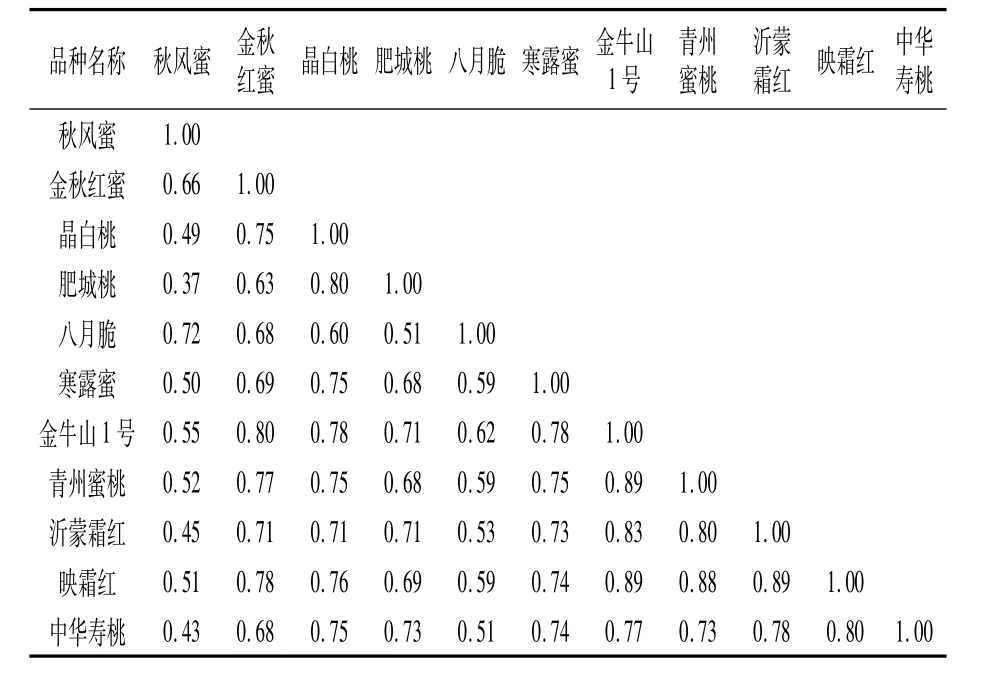
表4 11個桃品種間的遺傳相似系數
根據遺傳相似系數對桃品種進行主坐標分析,并繪制二維散點平面分布圖。如圖3所示,第一主坐標和第二主坐標分別解釋了32.85%和15.73%的品種間相關性,累計貢獻率為48.58%,能代表原始數據的主要信息。主坐標分析圖中,種質材料間的距離越近,表示親緣關系越近,反之則越遠。根據種質材料間的距離可將11份桃品種分成A、B和C三組,圖中分別用虛線圈出,進一步分析發現A、B組和C組與聚類分析結果(Ⅰ、Ⅱ組和Ⅲ組)高度相似,但主坐標分析更能直觀清楚地顯示桃品種間的親緣關系。
3 討論與結論
桃品種選育和優系創質的主要方式是基于實生苗選種、自然芽變選種和雜交群體選種。由于骨干親本的重復利用,桃種質材料的形態特征具有一定相似性,這為利用外觀形態特征進行辨別分類帶來一定困難。
DNA分子標記技術由于其穩定、高效、不易受外界環境影響的優點,已廣泛應用于果樹遺傳學分析,如芽變材料鑒定[9]、親緣關系分析[10-12]和種內亞群體劃分等[13]。SCoT分子標記具有高效、重復性好、應用性廣、簡單、成本低等優點,已經應用于茶樹[14]、楊桃[15]、月季[16]、枸杞[17]、龍眼[18]、柿[19]、荔 枝[20]等 物 種 中。本 研 究 利 用SCoT標記在桃中進行種質資源遺傳多樣性分析,發現14條引物在11個桃品種上共擴增出170條清晰、整齊的條帶,多態性水平較高(平均多態性比率為68.43%)。桃材料的遺傳相似系數在0.37~0.89之間,表明所收集的桃資源內具有較高的遺傳多樣性。根據聚類分析和主成分分析結果,11個桃品種可分為三大類群:第一類為中熟2個品種,第二類為晚熟與極晚熟青州蜜桃群組,第三類為晚熟肥城桃群組。Ⅰ組和Ⅱ、Ⅲ組間比較發現,品種多樣的遺傳背景與果實的不同成熟期具有一致性,表明二者緊密相關。Ⅱ組和Ⅲ組組間比較發現,同為晚熟桃品種,以青州蜜桃和肥城桃為代表的地方名優品種分組清晰明確,說明兩個組間具有成熟期之外的遺傳物質多樣性。
因此,利用SCoT標記對桃種質資源進行鑒定分析是十分有效的,具有較高的多態性檢測率,適用于桃種質資源鑒別及親緣關系分析,為桃資源的分類和種質的創制提供了科學依據。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
