谷子bZIP轉錄因子家族的全基因組鑒定與生物信息學分析
2020-09-14 08:19:38盧平武懿茂武強強李雪垠
山西農業科學 2020年9期
關鍵詞:植物
盧平,武懿茂,武強強,李雪垠
(1.山西農業大學生命科學學院,山西太谷030801;2.山西農業大學農學院,山西太谷030801)
谷子(Setaria italica)屬于1年生草本植物,為二倍體禾本科作物(2n=18),起源于我國,具有悠久的栽培歷史,我國的黃河中上游為主要栽培區。谷子是C4植物,耐旱、耐貧瘠,是發掘農作物抗旱基因和研究作物抗旱分子機制的優良種質資源[1]。2012年谷子的基因組測序已經完成,為谷子功能基因組學研究提供了數據平臺[2-3]。目前,谷子逐漸成為理想的模式作物之一,主要應用于禾本科植物的基因組學研究。谷子脫殼后稱為小米,小米富含蛋白質、脂肪、鐵、類胡蘿卜素、纖維素和維生素等,其營養價值很高;還可作為藥物[4],具有清熱、滋陰、治水瀉等功效;又可以用來釀酒。谷子的莖葉又可以用作牲畜的優等飼料。因此,其越來越受到人們的重視。
轉錄因子能調控下游基因的時空特異性表達,是真核生物中重要的調控蛋白。在植物中,轉錄因子是調節生長發育和脅迫應答過程的關鍵因子。堿性亮氨酸拉鏈(Basic leucine zipper,bZIP)轉錄因子廣泛分布于真核生物基因組,是數量最大、最保守的基因家族之一,在植物的生長發育、病原防御、高溫以干旱脅迫應答等生物和非生物脅迫反應中起著重要的調控作用[5]。因此,bZIP轉錄因子的基因鑒定和功能分析等研究在后基因組時代中是較為活躍的研究領域。bZIP轉錄因子是以共有的bZIP保守結構域為依據而被命名的。bZIP結構域包括一個C端的堿性氨基酸區域和一個N端的亮氨酸拉鏈區域,bZIP保守結構域包含了60~80個氨基酸,其中,堿性區域含有一個保守基序N-x7-R/K-x9,該保守基序由16~20個氨基酸殘基組成,這個區域參與細胞核的定位與DNA識別[6-7];亮氨酸拉鏈結構域由若干個兩親α螺旋構成,在α螺旋中每隔6個氨基酸就出現一個亮氨酸或其他疏水性的氨基酸,例如纈氨酸、甲硫氨酸、異亮氨酸和苯丙氨酸。
在植物中,bZIP轉錄因子調節包括病原防御、光反應和脅迫信號、種子成熟和花發育等過程[8]。在很多植物中,bZIP轉錄因子家族成員已經被鑒定或預測完成。在模式植物擬南芥和水稻中,分別包含75個和89個bZIP家族的成員[9-10]。在擬南芥中,bZIP轉錄因子能夠通過ABA途徑調節植物生長發育和非生物脅迫等過程[11];在水稻中,bZIP轉錄因子與干旱脅迫和花粉發育相關[12-13];在小麥中,有48個bZIP基因與花藥發育有關[14];在辣椒中,bZIP轉錄因子能夠抵御病原感染和非生物脅迫[15]。迄今為止,bZIP轉錄因子在擬南芥和水稻中的抗逆性研究較為深入,但在谷子中的研究卻相對較少[16]。
本研究利用生物信息學方法對谷子bZIP轉錄因子進行全基因組鑒定,并分析其家族成員的系統進化關系、基因結構、蛋白質理化性質、保守基序、啟動子順式作用元件和表達量水平,旨在為進一步研究谷子bZIP基因功能和分子機制奠定理論基礎。
1 材料和方法
1.1 全基因組鑒定和蛋白質理化性質預測
根據Pfam(http://pfam.xfam.org/)數據庫中谷子bZIP轉錄因子的Pfam編號,在Phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)v12數據庫中篩選出85個bZIP轉錄因子基因,并下載它們的基因組序列、編碼區序列(Coding sequence,CDS)和蛋白質序列。根據bZIP轉錄因子在染色體上的位置命名,谷子bZIP轉錄因子基因的染色體定位和基因組位置信息來源于Phytozome。利用ExPASy-Compute pI/Mw(https://web.expasy.org/compute_pi/)網站計算谷子bZIP轉錄因子蛋白質的理論等電點和分子量。
1.2 系統進化和基因結構預測
利用MEGA 7.0軟件對85條谷子bZIP轉錄因子的氨基酸序列進行多序列比對(Clustal W);采用Neighbor-Joining法構建系統發育樹,參數Bootstrap值設為1 000,其余參數默認。利用Gene Structure Display Server 2.0(http://gsds.cbi.pku.edu.cn/)網 站預測谷子bZIP轉錄因子基因結構,包括外顯子、內含子和上下游區域。
1.3 蛋白質模體和基因啟動子順式作用元件預測
利用網站MEME(http://meme-suite.org/)上傳氨基酸序列,分析谷子bZIP轉錄因子的蛋白質模體,模體數目設置為15個。
利用Phytozome網站的數據庫Setaria italica v2.2下載bZIP轉錄因子基因的5′UTR上游1.5 kb序列(包括5′UTR)作為谷子bZIP轉錄因子基因啟動子序列。利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)網站在線預測谷子bZIP轉錄因子基因啟動子上的順式作用元件。
1.4 基因組織特異性和環境誘導表達預測
利用TBtools,根據谷子bZIP轉錄因子基因在不同組織中和不同環境中的表達數據繪制熱圖。谷子bZIP轉錄因子基因不同組織、不同條件下的表達量數據來源于Phytozome Setaria italica v2.2數據庫。
2 結果與分析
2.1 全基因組鑒定及其蛋白質基本理化性質分析
表1對谷子bZIP轉錄因子的基因編號、染色體定位、基因組位置、基因長度、CDS長度、氨基酸個數、蛋白質的理論等電點和分子量進行了統計,利用Phytozome網站在谷子全基因組水平鑒定了bZIP轉錄因子基因共85個,根據bZIP轉錄因子在染色體和基因組位置上的分布,按順序依次命名為SibZIP1~SibZIP85。谷子bZIP轉錄因子基因在1~9號染色體上的數量分別是10、15、13、10、12、7、4、3、11個,分布不均勻。

表1谷子bZIP轉錄因子基因基本信息及其蛋白質基本理化性質

續表1
在谷子bZIP轉錄因子的蛋白質中,氨基酸個數最多的蛋白質是SibZIP35,有645個;最少的是SibZIP52,有140個。谷子bZIP轉錄因子家族蛋白質的理論等電點介于4.69~11.97,分子量在15.507~68.096 ku。蛋白質等電點偏高通常與其較高的堿性氨基酸(如精氨酸、賴氨酸、組氨酸)含量有關。例如SibZIP38、SibZIP61、SibZIP78、SibZIP83蛋白質等電點較高,分別為11.41、11.97、11.03、10.17,因為這幾個SibZIP轉錄因子蛋白質有較多的堿性氨基酸,因此等電點也較高。
2.2 系統進化樹的構建和基因結構分析
對谷子85條bZIP轉錄因子的氨基酸序列進行多序列比對,并構建系統進化樹如圖1(左)所示,部分SibZIP轉錄因子家族成員親緣關系較為相近,如SibZIP19/SibZIP68、SibZIP85/SibZIP79、SibZIP74/SibZIP70、SibZIP23/SibZIP80、SibZIP1/SibZIP51、SibZIP35/SibZIP22、SibZIP4/SibZIP44、SibZIP33/SibZIP60、SibZIP34/SibZIP48、SibZIP74/SibZIP70。因此,它們可能是旁系同源基因,由同一個bZIP轉錄因子祖先基因復制產生。圖1(右)為谷子bZIP轉錄因子基因的外顯子-內含子的結構情況分析。由圖1可知,85個谷子bZIP轉錄因子中親緣關系較為相近的基因結構也較為相似,如SibZIP19/SibZIP68、SibZIP85/SibZIP79、SibZIP74/SibZIP70、SibZIP23/SibZIP80,每組外顯子的個數都相等、長度相似,說明親緣關系較近的SibZIP基因的基因結構具有較強的保守性。
2.3 蛋白質模體和啟動子順式作用元件分析
利用網站MEME分析谷子bZIP轉錄因子蛋白質模體,結果如圖2所示,谷子bZIP轉錄因子蛋白質預測了15個模體,分別為模體motif1~motif15;85個bZIP蛋白質都含有motif1,說明motif1在谷子bZIP蛋白質中分布最廣,保守性最強;motif1有41個氨基酸,其中,5、10、17、19位的R(精氨酸)和27、34、41位的L(亮氨酸)具有較強的保守性,符合bZIP堿性亮氨酸拉鏈的特征。
部分bZIP轉錄因子蛋白質模體差異較大,一些親緣關系較近的bZIP轉錄因子蛋白質模體也較為相似。如SibZIP23和SibZIP80都具有motif1、motif4和motif12這3個motif;SibZIP2和SibZIP50、SibZIP4和SibZIP44、SibZIP34和SibZIP48都僅有motif1;SibZIP25和SibZIP81、SibZIP57和SibZIP31都具有motif1、motif8、motif10、motif11。
bZIP轉錄因子在植物中參與生長發育、光的形態建成和防御與脅迫應答等生理過程。為了探究SibZIP轉錄因子基因啟動子區域中順式作用元件與谷子生理過程的關系,本研究利用PlantCARE網站分析85個bZIP基因的啟動子順式作用元件,部分結果如表2所示,其中,AuxRR-core、TGA-element和P-box元件功能涉及植物生長發育過程的激素調控;G-box、GT1-motif和Sp1元件功能涉及植物光反應調控;ABRE元件功能涉及植物ABA響應。ABA不僅能夠抑制植物生長,還能夠在干旱、寒冷、高溫、鹽漬和水澇等逆境中增強植物抗逆性。MBS元件是MYB結合位點,其功能涉及植物干旱誘導;TC-rich repeats元件功能涉及植物防御和應激反應;ABRE、MBS和TC-rich repeats元件涉及防御與脅迫應答。
SibZIP78基因啟動子中含有16個G-box元件;SibZIP75基因啟動子中含有11個G-box元件和1個Sp1元件;SibZIP78、SibZIP54基因啟動子區域內含有較多的光響應相關順式作用元件;SibZIP54基因啟動子中含有1個AuxRR-core元件和3個TGA-element元件,SibZIP79基因啟動子中含有1個AuxRR-core元件、1個P-box元件和1個TGA-element元件,SibZIP54、SibZIP79基因啟動子富含激素調控相關的順式作用元件;SibZIP78基因啟動子中含有15個ABRE元件,SibZIP71基因啟動子中含有14個ABRE元件,SibZIP2基因啟動子中含有12個ABRE元件,SibZIP73基因啟動子中含有9個ABRE元件和2個MBS元件,這4個基因啟動子富含ABRE元件或MBS元件等與脅迫應答相關的順式作用元件(表2、3)。
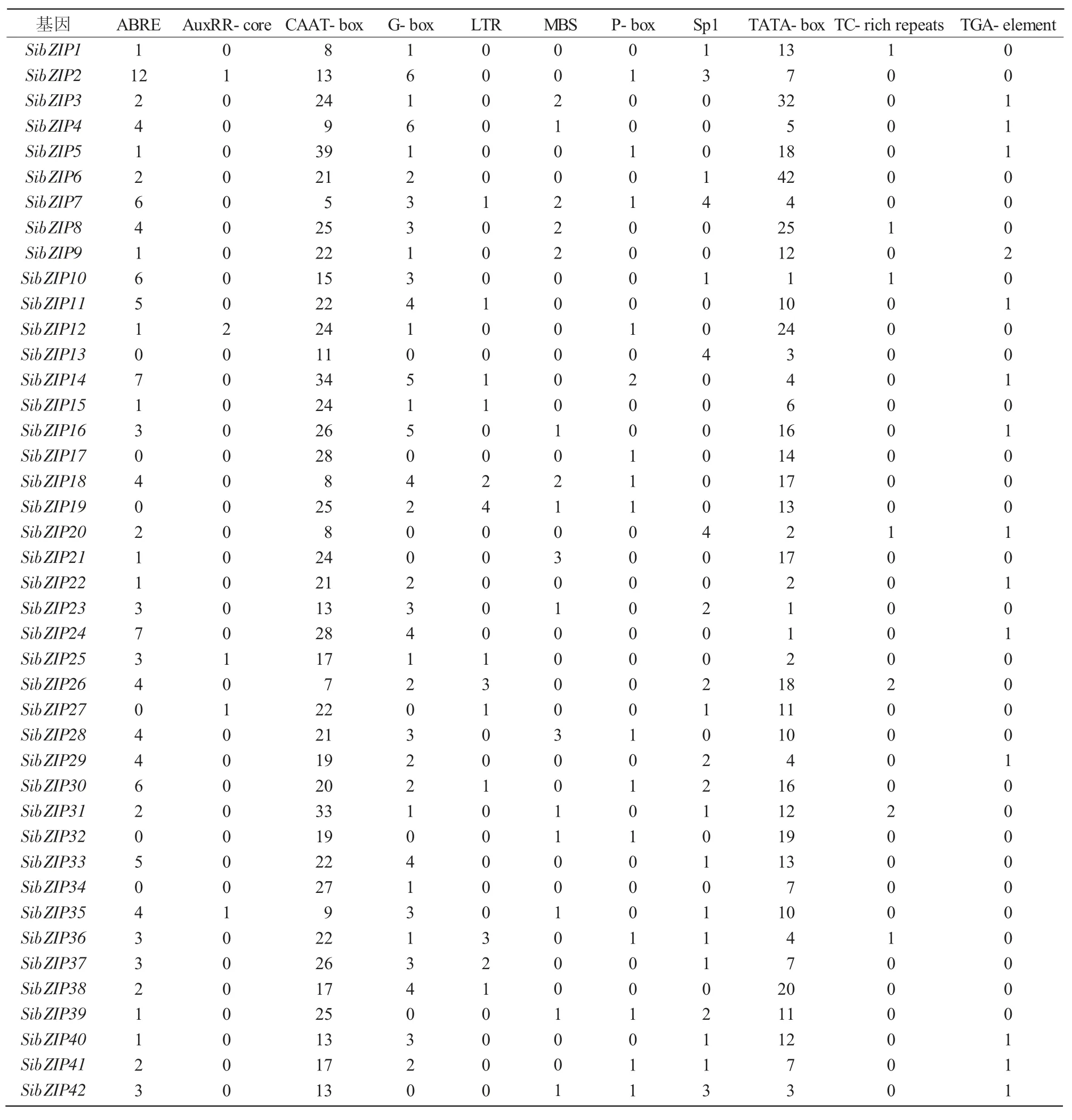
表2谷子bZIP基因啟動子順式作用元件種類與數目
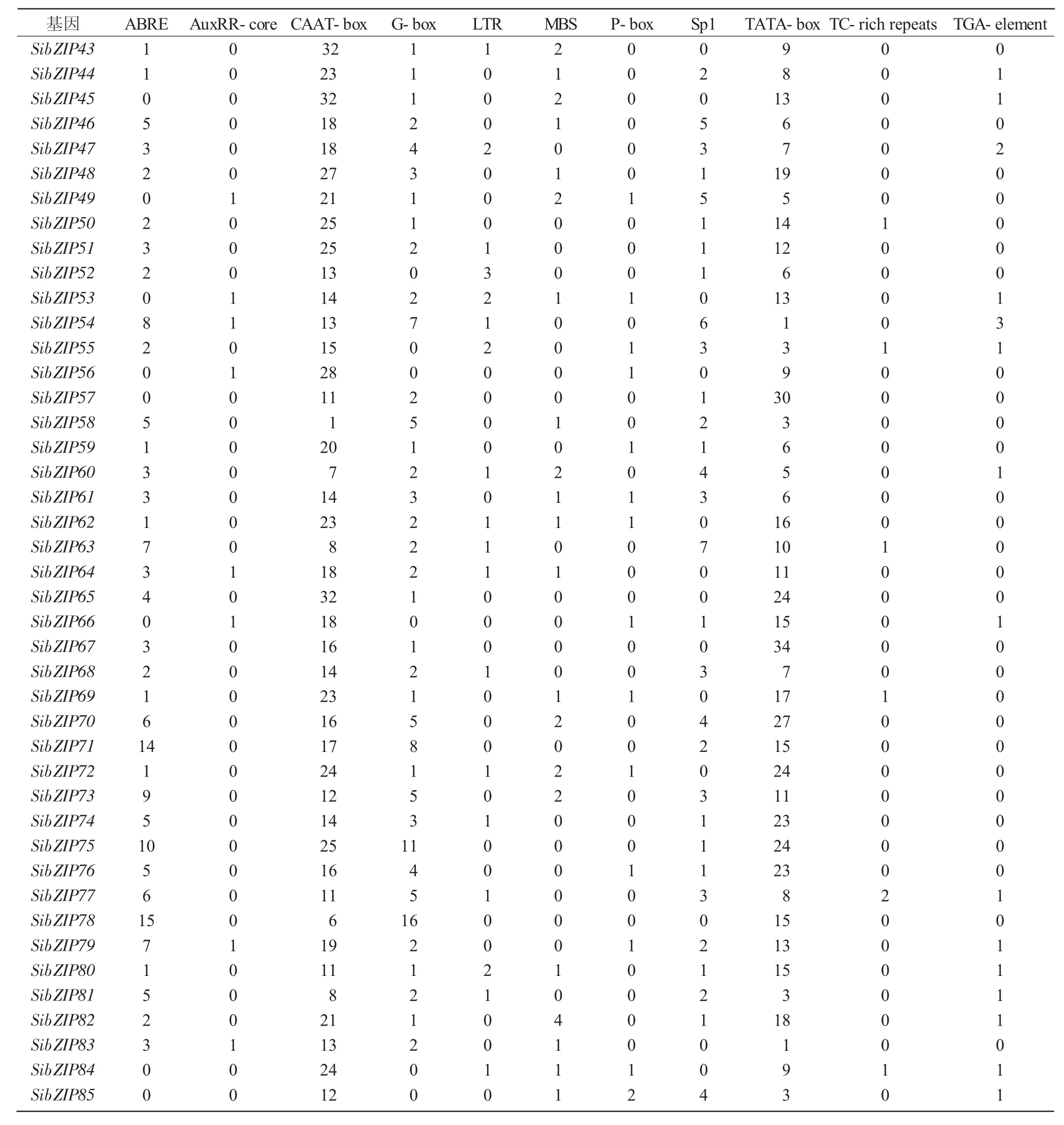
續表2

表3谷子bZIP基因啟動子順式作用元件核心序列與功能
2.4 組織特異性和環境誘導表達分析
利用TBtools軟件制作熱圖,可以較為明顯地顯示出谷子bZIP轉錄因子基因在不同組織及不同環境誘導的表達情況(圖3),表達情況相似的基因聚在一起。SibZIP3、SibZIP14、SibZIP48、SibZIP66等在不同條件下(10 d光照處理、干旱處理、硝酸處理、尿素處理、氨處理)的根中均有較高的表達量;SibZIP39、SibZIP75和SibZIP76等在圓錐花序中具有較高的表達量;SibZIP19、SibZIP20和SibZIP46等主要在光照下的葉片中表達;SibZIP21、SibZIP67、SibZIP79和SibZIP85均在黃化苗(Etiolated seedling)暗處理培養5 d的情況下具有較高的表達量;SibZIP31在遠紅光處理下特異表達;SibZIP17和SibZIP71在暗處理的地上部分中表達量較高;SibZIP11、SibZIP12和SibZIP72等在干旱處理的根中表達量較高。
3 結論與討論
谷子是C4植物,具有耐旱、耐貧瘠、水分利用率高和蒸騰系數低等特性,對生長環境要求較低,耐逆境脅迫能力較其他作物更強。因此,對谷子的基礎研究和抗逆基因資源的挖掘成為新的研究熱點。bZIP轉錄因子在植物中不僅能夠調節病原防御、脅迫應答等過程,還能夠調控光的形態建成、激素信號傳導、花發育和種子成熟與萌發等過程。植物的抗逆境脅迫反應機制非常復雜,因此參與調控植物抗逆脅迫反應的轉錄因子很多,除bZIP外,還有MYB、NAC、WRKY、HSF等[16-21]。
本研究利用生物信息學手段對谷子基因組中bZIP轉錄因子進行全基因組鑒定,并對所有家族成員進行了系統的生物信息學分析,經鑒定得到的85個SibZIP轉錄因子,分布在谷子的第1~9條染色體上,部分SibZIP轉錄因子家族成員具有一定保守性。親緣關系相近的轉錄因子往往是由具有相似的基因結構和相似蛋白質模體組成,如SibZIP4/SibZIP44、SibZIP34/SibZIP48。SibZIP轉錄因子家族保守基序較多、較為復雜,并且SibZIP轉錄因子具有多種多樣的生物學功能。SibZIP轉錄因子家族成員基因啟動子順式作用元件種類較多,包括與谷子防御脅迫相關的順式作用元件和谷子生長發育過程相關的順式作用元件。如SibZIP78、SibZIP75啟動子區域內含有較多與光響應有關的順式作用元件。因此,這2個基因可能參與谷子光反應調控。SibZIP54、SibZIP79基因啟動子中含有AuxRR-core、P-box或TGA-element元件的SibZIP轉錄因子,可能參與植物生長素、赤霉素的調控;SibZIP2、SibZIP71、SibZIP73等轉錄因子基因啟動子富含ABRE、MBS、TC-rich repeats或LTR元件的SibZIP轉錄因子,可能參與植物非生物脅迫調節、防御和應激反應。
通過對SibZIP轉錄因子家族成員表達量分析可知,SibZIP3/SibZIP14/SibZIP48、SibZIP39/SibZIP75/SibZIP76和SibZIP19/SibZIP20/SibZIP46分別在根、圓錐花序和葉片中具有較高的表達量。因此,推測這些bZIP轉錄因子在根、圓錐花序和葉片中特異表達,參與根、圓錐花序和葉片的生長發育。SibZIP21和SibZIP67均在黃化苗暗處理情況下具有較高的表達量,并且SibZIP21和SibZIP67基因結構和蛋白質模體較為相似。因此,推測它們功能相似,可能參與谷子暗反應。SibZIP30和SibZIP61在光照處理的根中具有較高表達量,說明這2個SibZIP基因可能參與谷子根部光反應。SibZIP31在遠紅光處理下特異表達,說明遠紅光能誘導SibZIP31基因表達。SibZIP17和SibZIP71在暗處理地上部分中表達量較高,說明黑暗能誘導SibZIP17和SibZIP71表達。SibZIP11、SibZIP12和SibZIP72等在干旱處理的根中表達量較高,說明干旱能夠誘導其表達,這3個SibZIP基因可能參與谷子根部的干旱脅迫應答。
在植物中,bZIP轉錄因子的功能是多樣的,不同的家族成員作用不同。因此,對bZIP轉錄因子的功能和作用機制的研究十分重要。高等植物中部分bZIP轉錄因子家族成員已經有了較為廣泛的研究,接下來可以利用基因克隆、蛋白質組學等技術進一步研究bZIP轉錄因子在植物中的調控機制,從而更深入了解高等植物遺傳進化、生長發育和脅迫應答的過程。
本研究依據生物信息學方法,對谷子bZIP轉錄因子進行全基因組鑒定,對基因結構、蛋白質理化性質、蛋白質模體、啟動子順式作用元件和表達量水平進行了預測分析,其結果為更深入研究谷子bZIP轉錄因子的生物學功能及其在谷子中的生長發育調控和逆境脅迫響應機制提供了參考。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
