蘋果屬種質TP-M13-SSR親緣關系及遺傳多樣性分析
2020-09-14 08:19:40侯麗媛董艷輝聶園軍張春芬李亞莉王育川曹秋芬
山西農業科學 2020年9期
侯麗媛,董艷輝,聶園軍,鄧 舒,肖 蓉,張春芬,李亞莉,趙 菁,王育川,曹秋芬
(1.山西農業大學生命科學學院,山西太原030031;2.山西農業大學農業經濟管理學院(山西省農業科學院農業資源與經濟研究所),山西太原030006;3.山西農業大學果樹研究所,山西太原030031)
蘋果是世界上最重要的果樹之一。對蘋果種質資源的準確鑒定是蘋果種質資源保存和利用的前提。蘋果種質資源的鑒定方法有多種,如形態學觀察、同工酶[1-2]以及分子標記[3-5]等。而SSR分子標記即簡單重復序列(SSR),又稱微衛星標記,是普遍存在且隨機分布于真核生物基因組中的一種特殊序列,SSR標記具有共顯性、豐富的多態性、高度重復性、高度可靠性以及操作簡單等優點,是近年來應用廣泛的一類特異性強的分子標記。
TP-M13-SSR(Simple sequence repeat with tailed primer M13)技術是熒光測序技術和SSR技術相結合形成的一種對擴增產物進行熒光檢測的體系[6],該技術具有結果重復性高、準確性好的特點,并在較大程度上解決了擴增產物檢測流程繁瑣、分析通量較低、記錄數據工作量過大等一系列問題[7]。目前,TP-M13-SSR技術已經被應用到許多作物的種質資源研究中。張志軍等[8]利用TP-M13-SSR標記技術對23個麥芽品種進行了指紋圖譜構建。樊云芳等[9]運用TP-M13-SSR技術對29份枸杞種質資源進行了遺傳多樣性分析。BARKLEY等[10]對花生的遺傳多樣性進行了評價。有關果樹方面,高源等[11]構建了我國原產蘋果屬27份材料在12個SSR位點的指紋圖譜;CAO等[12]對我國72份野生山梨進行了遺傳多樣性分析。
本研究利用16對TP-M13-SSR標記對山荊子和24份我國華北地區部分蘋果栽培品種的指紋圖譜進行構建,并在此基礎上對試驗材料進行親緣關系和遺傳多樣性分析,旨在為后期更有效地利用這些蘋果種質資源進行研究提供重要依據。
1 材料和方法
1.1 材料
選取目前我國華北地區部分蘋果屬25份材料,其中,蘋果系材料24份,砧木系材料1份(表1),25份材料均取自國家果樹種質興城蘋果資源圃(中國農業科學院果樹研究所)。
1.2 試驗方法及數據處理
1.2.1 試驗方法 采用德國QIAGEN的DNeasy Plant Mini Kit試劑盒分別提取各供試材料春季嫩葉基因組DNA。選取245對擴增片段長度在100~300 bp的普通SSR序列,參照YAMAMOTO等[13]、GIANFRANCESCHIá等[14]、LIEBHARD等[15]和高源等[16]的研究,共合成16對TP-M13-SSR引物。TPM13-SSR引物是由3條引物組成,其中,第1條引物是普通SSR引物的正向引物;第2條引物由帶有M13接頭的正向引物和普通SSR引物的反向引物相連而成;第3條引物是5′端帶有熒光標記的M13正向引物。上海Sangon公司合成第1、2條引物;美國ABI公司合成第3條引物,5′端帶有熒光標記的M13正向引物標記的3種熒光分別為6FAMTM(Blue)、VICTM(Green)和NEDTM(Yellow)(表2)。

表2第3條引物及序列
1.2.2 PCR擴增和純化體系16對TP-M13-SSR 引物的名稱、序列及優化退火溫度如表3所示。

表3 16對TP-M13-SSR引物序列及其優化條件
1.2.2.1 試驗程序[5,11-12]因TP-M13-SSR引物的2條序列之間存在19 bp的差別,因此,TP-M13-SSR引物的PCR反應需要分2步進行,第1步需要獲得TP-M13-SSR引物的擴增產物;第2步是實現SSR擴增產物的熒光標記,即5′端帶有熒光標記的M13正向引物的擴增。
TP-M13-SSR引物PCR反應體系為:DNA(20 mg/L)2μL,F-primer(2μmol/L)0.12μL,Rprimer(2μmol/L)1.2μL,10×PCR buffer(不含Mg2+)1μL,Mg2+(25 mmol/L)0.8μL,dNTP(25 mmol/L)0.1μL,Taq酶(5 U/μL)0.12μL,ddH2O補足10μL。PCR反應程序為:94℃預變性5 min;94℃變性45 s,退火(根據不同引物選擇不同的退火溫度)45 s,72℃延伸45 s,35個循環;72℃保持10 min。4℃保存備用。
以TP-M13-SSR引物PCR產物為模板,加入5′端帶有熒光標記的M13正向引物(2μmol/L)1.8μL,10×PCR buffer(不含Mg2+)0.1μL,Taq酶(5 U/μL)0.08μL,ddH2O 0.12μL。PCR反應程序:94℃預變性5 min;94℃變性30 s,53℃退火30 s,72℃延伸30 s,16個循環;72℃保持10 min。4℃保存備用。1.2.2.2 PCR產物的純化 根據熒光標記引物的不同顏色以及擴增產物分子量的大小進行混合純化。取2μLPCR產物,在其中加入無水乙醇6μL,輕輕振蕩2~3 min混勻,然后3 000 r/min離心30 min;輕輕倒出液體,倒置離心管片刻后于700 r/min離心30~60 s,甩干液體;加入70% 乙醇60μL,輕輕振蕩2~3 min,3 000 r/min離心10 min;輕輕倒出液體,倒置離心管片刻后于700 r/min離心30~60 s,靜置5 min;加入60μL ddH2O,振蕩混勻,室溫避光溶解1 h。4℃保存備用。
用測序儀ABI3730對純化的TP-M13-SSR引物的PCR產物進行自動熒光檢測,并對獲得的不同樣品在每個TP-M13-SSR位點的擴增片段長度進行收集。
1.2.3 數據處理 將收集到的原始數據轉換成以1和0標記的數據值矩陣,采用NTSYS-pc 2.10e分析軟件,用Jaccard相似系數進行計算,得到相似系數矩陣。在此基礎上,采用UPGMA(Unweighted Pair-Group Method with Arithmetic Mean)方法對試驗材料進行聚類分析,得到這些材料的親緣關系樹狀圖。最后利用GenAlEx 6.501軟件對供試材料進行遺傳多樣性分析。
2 結果與分析
2.1 25份試材的指紋圖譜分析
利用16對TP-M13-SSR引物對25份蘋果屬材料進行PCR檢測,準確獲得了不同材料在16個TP-M13-SSR位點的指紋信息(表4)。以種植面積相對較大的蘋果品種最良富士為例進行指紋圖譜分析。最良富士在4對TP-M13-SSR引物位點的毛細管電泳圖譜如圖1所示,在每個TP-M13-SSR位點獲得的等位基因數即為表4中最良富士在該位點對應的擴增片段的長度。
每對TP-M13-SSR引物的擴增帶型為3~13個,平均為8.716個。25份蘋果屬供試材料在16對TP-M13-SSR引物位點獲得的指紋數據互不相同,該指紋數據即為各供試材料的指紋圖譜,這些指紋圖譜也是各品種的特定譜帶,該譜帶可以作為鑒定各品種的依據。

表4 25份蘋果屬種質資源在16對TP-M13-SSR引物位點的指紋圖譜
2.2 擴增產物的多態性分析
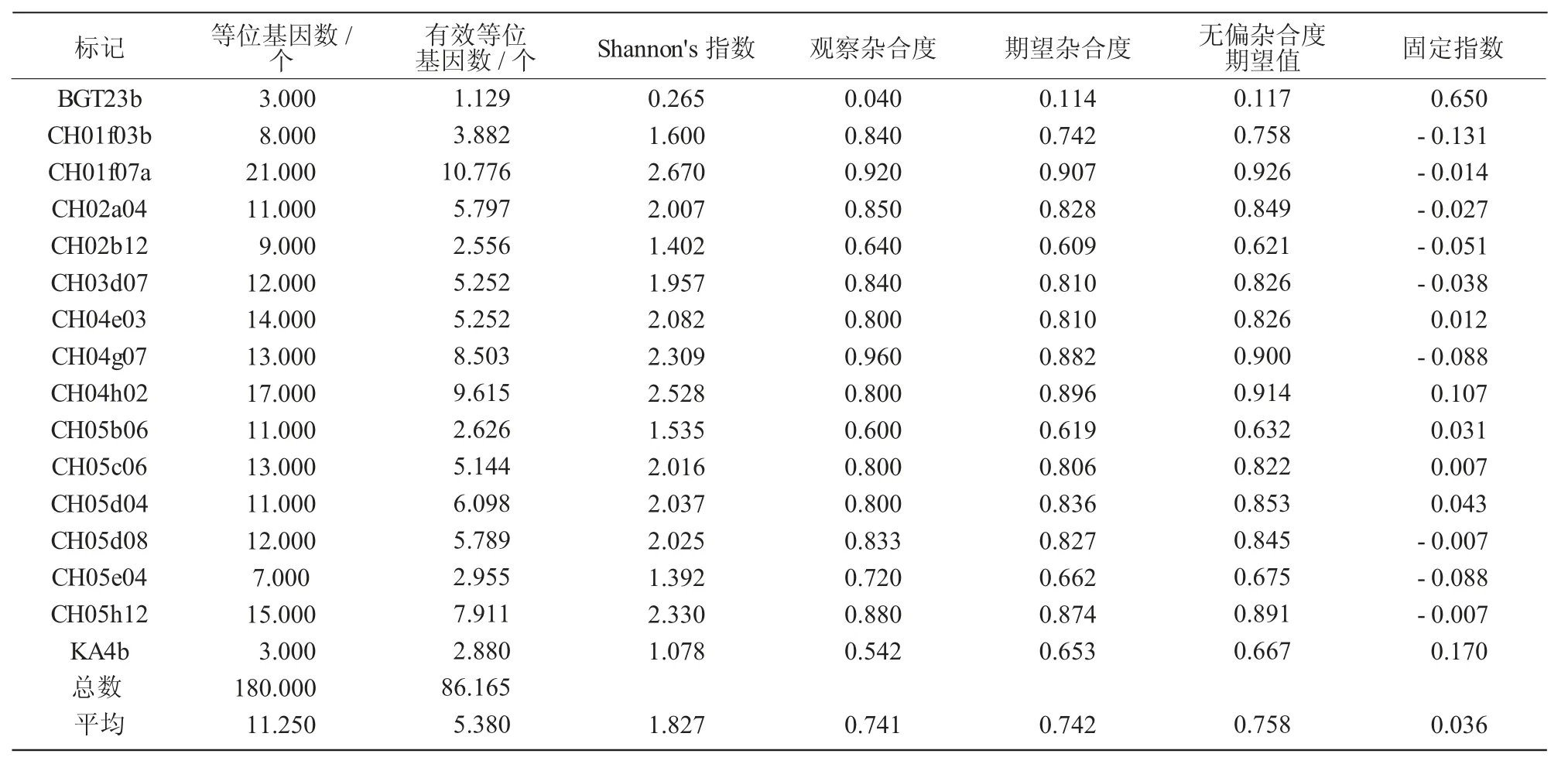
表5 25個蘋果屬種質在16對TP-M13-SSR引物位點檢測到的的遺傳多樣性
由表5可知,供試的25份蘋果屬種質在16對TP-M13-SSR引物中共擴增出等位基因180個,平均每對TP-M13-SSR引物觀測到的等位基因數為11.25個,有效等位基因數為5.38個;不同供試材料在每個TP-M13-SSR位點檢測到的等位基因數不同,3~21個不等,其中,在SSR位點BGT23b和KA4b檢測到等位基因數為3個,在SSR位點CH01f07a檢測到等位基因數為21個。擴增片段長度大小范圍主要在104~245 bp,其中,SSR引物CH04h02的擴增片段大小范圍最大,為155~245 bp,相差90 bp;引物KA4b擴增片段大小范圍最小,為155~159 bp,相差4 bp。Shannon's指數從BGT23b的0.265到CH01f07a的2.670,平均為1.827。等位基因數和Shannon's指數均高于平均值的SSR引物有CH03d07、CH04e03、CH04h02、CH05c06、CH05d08、CH01f07a、CH04g07、CH05h12。觀測雜合度為0.040(BGT23b)~0.096(CH04g07),平均為0.741;期望雜合度值從BGT23b的0.114到CH01f07a的0.907,平均為0.742;無偏雜合度期望值從BGT23b的0.117到CH01f07a的0.926,平均為0.758。固定指數平均為0.036,說明供試蘋果品種間含有的純合子較多。
2.3 親緣關系分析
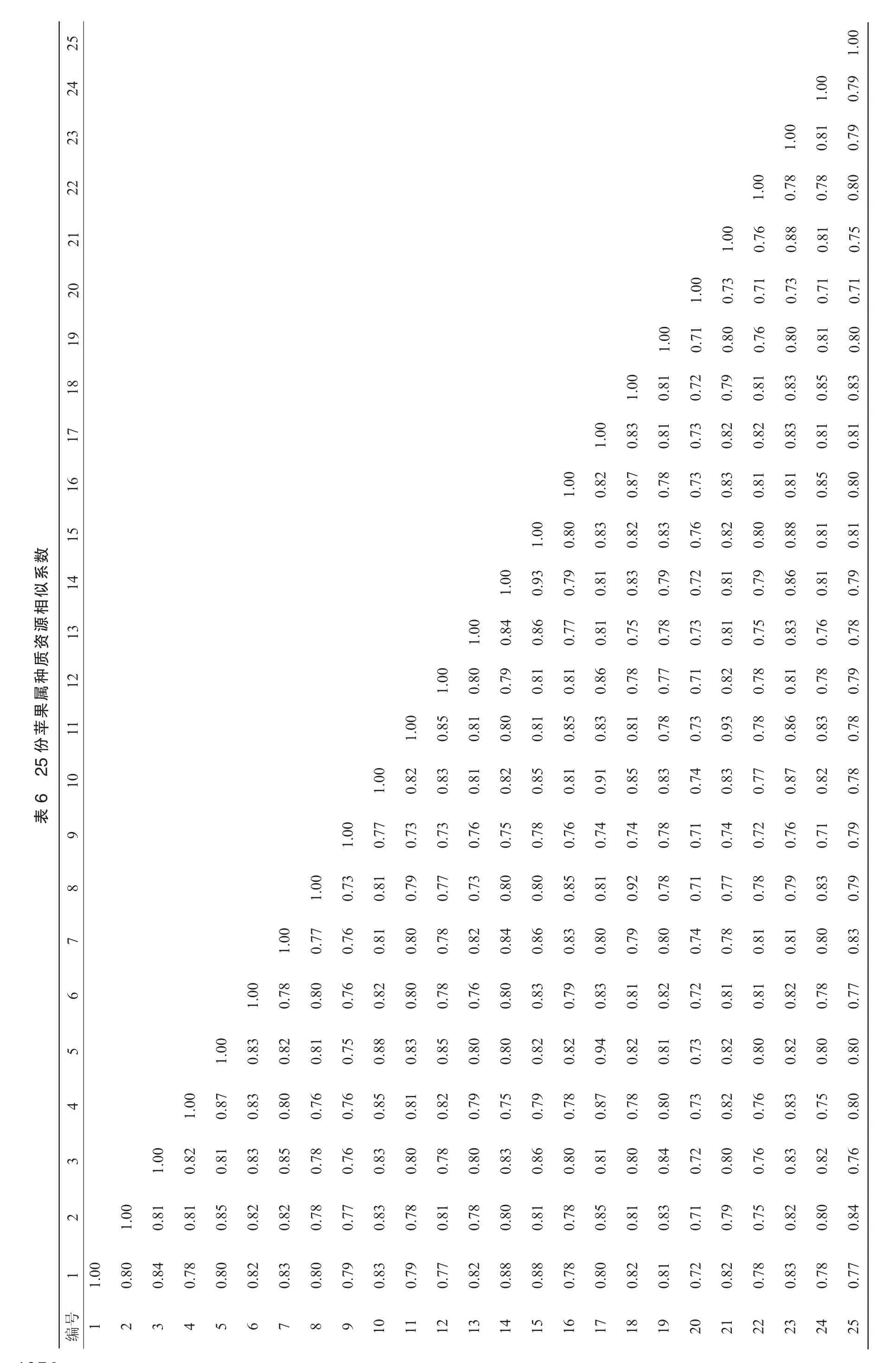
根據16對TP-M13-SSR引物的擴增數據,采用NTSYSpc 2.10e軟件計算不同種質之間的相似系數,結果表明(表6),25份蘋果屬材料的相似系數在0.72~0.94,平均達到0.83,說明在分子水平上各材料間的遺傳親緣關系較近。
25 1.00 24 1.00 0.79 23 1.00 0.81 0.79 22 1.00 0.78 0.78 0.80 21 1.00 0.76 0.88 0.81 0.75 20 1.00 0.73 0.71 0.73 0.71 0.71 19 1.00 0.71 0.80 0.76 0.80 0.81 0.80 18 1.00 0.81 0.72 0.79 0.81 0.83 0.85 0.83 17 1.00 0.83 0.81 0.73 0.82 0.82 0.83 0.81 0.81 16 1.00 0.82 0.87 0.78 0.73 0.83 0.81 0.81 0.85 0.80數系似15 1.00 14 1.00 0.93 0.80 0.79 0.83 0.81 0.82 0.83 0.83 0.79 0.76 0.72 0.82 0.81 0.80 0.79 0.88 0.86 0.81 0.81 0.81 0.79相源資13 1.00 0.84 0.86 0.77 0.81 0.75 0.78 0.73 0.81 0.75 0.83 0.76 0.78質種屬12 1.00 0.80 0.79 0.81 0.81 0.86 0.78 0.77 0.71 0.82 0.78 0.81 0.78 0.79果蘋份25 6 11 1.00 10 1.00 0.82 0.85 0.83 0.81 0.81 0.80 0.82 0.81 0.85 0.85 0.81 0.83 0.91 0.81 0.85 0.78 0.83 0.73 0.74 0.93 0.83 0.78 0.77 0.86 0.87 0.83 0.82 0.78 0.78表9 1.00 0.77 0.73 0.73 0.76 0.75 0.78 0.76 0.74 0.74 0.78 0.71 0.74 0.72 0.76 0.71 0.79 8 1.00 0.73 0.81 0.79 0.77 0.73 0.80 0.80 0.85 0.81 0.92 0.78 0.71 0.77 0.78 0.79 0.83 0.79 7 1.00 0.77 0.76 0.81 0.80 0.78 0.82 0.84 0.86 0.83 0.80 0.79 0.80 0.74 0.78 0.81 0.81 0.80 0.83 6 1.00 0.78 0.80 0.76 0.82 0.80 0.78 0.76 0.80 0.83 0.79 0.83 0.81 0.82 0.72 0.81 0.81 0.82 0.78 0.77 5 1.00 0.83 0.82 0.81 0.75 0.88 0.83 0.85 0.80 0.80 0.82 0.82 0.94 0.82 0.81 0.73 0.82 0.80 0.82 0.80 0.80 4 1.00 0.87 0.83 0.80 0.76 0.76 0.85 0.81 0.82 0.79 0.75 0.79 0.78 0.87 0.78 0.80 0.73 0.82 0.76 0.83 0.75 0.80 3 2 1.00 1.00 0.81 0.82 0.81 0.81 0.85 0.83 0.82 0.85 0.82 0.78 0.78 0.76 0.77 0.83 0.83 0.80 0.78 0.78 0.81 0.80 0.78 0.83 0.80 0.86 0.81 0.80 0.78 0.81 0.85 0.80 0.81 0.84 0.83 0.72 0.71 0.80 0.79 0.76 0.75 0.83 0.82 0.82 0.80 0.76 0.84 1 1.00 0.80 0.84 0.78 0.80 0.82 0.83 0.80 0.79 0.83 0.79 0.77 0.82 0.88 0.88 0.78 0.80 0.82 0.81 0.72 0.82 0.78 0.83 0.78 0.77號1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25編
在相似系數0.72處,25份蘋果屬種質資源可以明顯分為2個類群,即山荊子組(Ⅰ)和蘋果組(Ⅱ),其中,山荊子組(Ⅰ),只有山荊子(20)1個種質,蘋果組(Ⅱ)包括其他24份蘋果栽培材料;在相似系數0.82處,蘋果組(II)又被分為8組(Ⅱ-1~II-8),其中,Ⅱ-1組中只有舞美(9)1個品種,Ⅱ-2組中也只有蜜脆(22)1個品種,Ⅱ-3組中有紅星(8)、元帥(18)、祝光(16)和青香蕉(24);Ⅱ-4組中是初秋(2)和旭(25);Ⅱ-5組中新嘎拉(6)和王林(19)聚在了一起;Ⅱ-6組有紅玉(11)、錦玉(21)和中秋(23);Ⅱ-7組中有金冠(5)、金矮生(17)、津輕(4)、丹霞(10)和喬納金(12);Ⅱ-8組中最良富士(1)、長富1號(14)、長富2號(15)、千秋(3)、國光(7)和寒富(13)聚在一起(圖2)。
3 討論
3.1 SSR引物的特點
SSR分子標記因其特性近年來得到了廣泛應用[17]。TP-M13-SSR技術是基于SSR分子標記技術發展起來的一種對擴增產物進行熒光檢測的體系,其實現了對SSR分子標記數據的準確和高效的收集,對僅有2 bp之差的差異片段能夠進行有效區別,非常適用蘋果種質資源指紋圖譜的構建[18-19]。本研究選用的16對TP-M13-SSR引物多態性較好,雜合度較高,在16對SSR引物位點獲得的指紋圖譜互不相同,可以作為各品種的特定譜帶,適宜構建蘋果品種SSR指紋圖譜及進行遺傳多樣性分析。
3.2 遺傳多樣性分析
觀測等位基因數越高則表明SSR位點的多態性和群體變異程度越高[20]。本研究對供試的25份蘋果屬種質在16對TP-M13-SSR引物位點進行擴增,共檢測到180個等位基因位點,平均每對引物觀測到的等位基因數為11.25個;每個TP-M13-SSR位點可檢測到的等位基因數為3~21個不等,大小范圍主要在104~245 bp,其中,在SSR位點BGT23b和KA4b檢測到等位基因數為3個,在SSR位點CH01f07a檢測到等位基因數為21個,擴增片段大小范圍主要在170~232 bp,SSR引物CH04h02的擴增片段大小范圍最大,為155~245 bp,相差90 bp;引物KA4b擴增片段大小范圍最小,為155~159 bp,相差4 bp。
平均有效等位基因數、平均Shannon信息指數、平均期望雜合度等是用來評價遺傳多樣性的常用指標。16對引物的平均有效等位基因數為5.38個,Shannon's指數從BGT23b的0.265到CH01f07a的2.670,平均為1.827,等位基因數和Shannon's指數均高于平均值的SSR引物有CH03d07、CH04e03、CH04h02、CH05c06、CH05d08、CH01f07a、CH04g07、CH05h12。觀測雜合度從BGT23b的0.040到CH04g07的0.096,平均為0.741;期望雜合度值從BGT23b的0.114到CH01f07a的0.907,平 均 為0.742;無偏雜合度期望值從BGT23b的0.117到CH01f07a的0.926,平均為0.758。
固定指數是群體中雜合子頻率差異的估計值,是通過雜合子頻率偏差來反映群體在某個座位的遺傳分化程度[21]。本研究中,固定指數在7對SSR引物位點為正值,平均為0.036,接近于0,說明供試蘋果品種間含有的純合子存在過量現象但是過量不明顯,也說明供試材料接近于Hardy-Weinberg平衡。鑒于此,在育種工作中要盡量擴大選擇范圍,引入新的種質,促使種群間的基因交流和重組,以提高后代的品質。
3.3 聚類分析
本研究根據16對TP-M13-SSR引物對供試的25份蘋果屬種質材料的擴增數據計算相似系數,并進行遺傳分析和聚類分析,結果顯示,25份材料的相似系數在0.72~0.94,平均達到0.83,說明在分子水平上各材料間的遺傳親緣關系較近。在相似系數0.72處,25份蘋果屬種質資源可以明顯分為2個類群,即山荊子組(I)和蘋果組(II),其中,山荊子組(I)只有山荊子(20)1個種質,蘋果組(II)包括其他24份蘋果栽培材料。種質間相似系數較低說明遺傳關系相對較遠,而相似系數較高則說明種質間具有相近的親本起源。本研究結果顯示,山荊子和其他蘋果栽培品種間遺傳相似系數相對較低,二者間親緣關系相對較遠,僅從遺傳相似系數的數值來說,山荊子和蘋果栽培品種間的遺傳分化不是太大。山荊子屬薔薇科蘋果屬,與蘋果嫁接后親和力強,是我國北方蘋果繁殖的主要砧木材料。本研究對山荊子作為蘋果常用砧木進行了分子水平的解釋。
在相似系數0.82處,蘋果組(II)又被分為8組(II-1~II-8),其中,II-1組中只有舞美(9)1個品種,舞美是英國東茂林試驗站從威賽克旭自然實生苗中選育的柱型蘋果,與其他栽培種蘋果親緣關系相對較遠;II-2組中也只有蜜脆(22)1個品種,蜜脆由Macoun×Honeygold雜交選育而來,原產于美國,說明蜜脆與其他蘋果品種之間遺傳關系相對較遠;II-3組中有紅星(8)、元帥(18)、祝光(16)和青香蕉(24),其中,紅星蘋果為元帥的芽變品種,祝光原產于美國,由實生選種而來,青香蕉由實生選種而來,說明這4個早期的蘋果實生品種的來源很相近,紅星(8)和元帥(18)在相似系數0.92處聚為一類,親緣關系較近,但與祝光(16)和青香蕉(24)親緣關系較遠;II-4組中有初秋(2)和旭(25),初秋是從紅玉和金冠的雜交種中選育而來,原產于日本,旭原產于加拿大,為實生選種,說明二者遺傳距離相對較近,初秋的親本紅玉和金冠與旭親緣關系較近;II-5組中新嘎拉(6)和王林(23)聚在了一起,新嘎拉是嘎拉((元帥×桔蘋)×金冠)的枝變,王林是由金冠×印度雜交選育而來,這2個品種更多地保留了金冠的遺傳特性;II-6組有紅玉(11)、錦玉(21)和中秋(23),其中,紅玉為可口香的實生后代,原產于美國,錦玉是紅玉的芽變,二者間遺傳距離較近,中秋是紅玉和元帥的雜交種,其更多保留了紅玉的遺傳特性;II-7組中有金冠(5)、金矮生(17)、津輕(4)、丹霞(10)和喬納金(12),其中,金矮生是金冠的短枝芽變,二者相似系數為0.94,親緣關系很近,津輕的母本為金冠,丹霞是由金冠實生苗選育而來,喬納金是金帥(金冠)×紅玉雜交種,說明津輕、丹霞和喬納金從遺傳學角度來說更多地保留了金冠的遺傳特性;II-8組中最良富士(1)、長富1號(14)、長富2號(15)、千秋(3)、國光(7)和寒富(13)聚在一起,其中,最良富士、長富1號、長富2號和寒富都為富士系,是富士芽變,國光為富士(國光×元帥)的親本,富士為千秋(東光×富士)的親本,這些品種都含有國光的血緣,表明從遺傳學上來講,富士及其后代保留了母本國光更多的遺傳特性。
4 結論
本研究結果顯示,TP-M13-SSR分子標記的分析結果與已知蘋果屬間的譜系較為一致,聚類分析結果能完全體現后代與親本之間的親緣關系,另外,從分子水平對山荊子作為蘋果常用砧木也進行了驗證。
