羅氏沼蝦轉錄組密碼子使用偏好性分析
2020-09-15 16:27:20慎佩晶張宇飛李喜蓮高強徐洋黃振遠蔣文枰陳雪峰
現代農業科技 2020年16期
慎佩晶 張宇飛 李喜蓮 高強 徐洋 黃振遠 蔣文枰 陳雪峰
摘要 ? ?以羅氏沼蝦轉錄組數據為數據來源,通過研究羅氏沼蝦轉錄組的密碼子使用參數(如有效密碼子的數量和相關密碼子堿基的具體組成信息等),并且采用Codon W 1.4.4深入開展了統計和計算。研究結果顯示,同義密碼子第三位核苷酸和表達基因密碼子GC含量均值分別為0.40和0.45。從整體上看,ENC的平均值等于52.72,其中絕大部分的ENC值小于35。采用高頻密碼子的研究方法獲得GAU、GAA、UUU、AAU、CCA等5個高頻密碼子。通過最優碼子分析法確定16個最優密碼子,編碼10個氨基酸,最優密碼子除UUG外均以A/T結尾。而且把它和大腸桿菌、酵母、果蠅以及人類等6種生物的密碼子使用頻率開展比較,結果表明,與大腸桿菌和果蠅存在較大差異,而與酵母最為接近。研究結果可為羅氏沼蝦功能基因和分子育種等提供理論基礎。
關鍵詞 ? ?羅氏沼蝦;偏好性;轉錄組;密碼子
中圖分類號 ? ?S968.12 ? ? ? ?文獻標識碼 ? ?A
文章編號 ? 1007-5739(2020)16-0175-04
Analysis of ?Codon ?Usage ?Bias ?Based ?on ?Macrobrachium ?rosenbergii ?Transcriptome
SHEN Pei-jing ? ?ZHANG Yu-fei ? ?LI Xi-lian ? ?GAO Qiang ? ?XU Yang ? ?HUANG Zhen-yuan ? ?JIANG Wen-ping ? ?CHEN Xue-feng *
(Genetics and Breeding Research Laboratory, Agriculture Ministry Key Laboratory of Healthy Freshwater Aquaculture, Key Laboratory of Freshwater Aquatic Animal Genetic and Breeding of Zhejiang province, Zhejiang Institute of Freshwater Fisheries,Huzhou Zhejiang 313001)
Abstract ? ?Using the transcriptome data of Macrobrachium rosenbergii as the data source, CodonW1.4.4 software was used to analyze and study the codon usage, and other codon usage parameters such as the transcript base composition and actual codon number of Macrobrachium rosenbergii, and the expression genes of Macrobrachium rosenbergii were found. The results showed that the average GC content of the codon was 0.45, and the average GC content of the third nucleotide of the synonym codon was 0.40. The mean ENC of the expressed gene was 52.72, and most of the ENC values were less than 35. Five high-frequency codons of GAU, GAA,UUU, AAU and CCA were determined by high-frequency codon analysis. Sixteen optimal codons were determined by optimal codon analysis and 10 amino acids were encoded. The optimal codons all ended in A/T except UUG. Comparing the codon usage frequency of Macrobrachium rosenbergii with six other species such as Escherichia coli, Saccharomyces cervisiae, Drosophila melanogaster, Homo sapiens, etc., it was found to be quite different from Escherichia coli and Drosophila melanogaster, but it was almost consistent with Saccharomyces cerevisiae. The research provided a theoretical basis for functional genome research and molecular breeding of Macrobrachium rosenbergii.
Key words ? ?Macrobrachium rosenbergii; usage bias; transcriptome; codon
羅氏沼蝦是我國重要的經濟型甲殼動物之一,目前,國內的羅氏沼蝦養殖規模已高居全球首位,2018年我國羅氏沼蝦苗種生產量達到3 000億尾,養殖面積大約14萬hm2,總產量60萬t。羅氏沼蝦主產區集中在浙江湖州、嘉興和杭州,江蘇高郵和江都,廣東高要、中山和珠海等地,近年來呈現向內陸及北方地區擴展的趨勢[1]。隨著高通量測序技術的進一步推廣,miRNA組學、轉錄組已知分子標記的開發已運用于羅氏沼蝦的高產、抗病等重要經濟性狀的研究。
作為信使RNA最重要的編碼單元之一,三聯密碼子所對應的氨基酸的翻譯起始或終止蛋白。研究團隊開展密碼子偏好性研究,通過分子標記的方式來進行輔助性育種工作,有助于確定物種間的進化關系,新基因的挖掘和其他生物學進一步使用,我國在該領域已開展數十年的研究[2]。當前密碼子的偏好性研究,在水生動物研究中得到廣泛應用。我國魚類研究大量使用了密碼子偏好性研究,例如團頭魴密碼子使用偏好性差異與親緣關系相一致,因而遺傳密碼子的偏好性能夠較好地反映鲌亞科魚類的進化關系,為探究屬內遺傳變異及密碼子優化和相關育種工作提供了分子材料[3]。金魚的密碼子偏好性與酵母的密碼子偏好性使用差異最大,而與斑馬魚的密碼子偏好性一致[4]。在貝類研究方面,由于優化了太平洋牡蠣 fut2基因的相關密碼子,從而為其特異性富集諾如病毒方面的基礎科學研究,尤其是分子生物學機理方面的探索打下了良好的理論基礎[5]。鰲蝦次目功能基因密碼子偏好性研究,為螯蝦次目新基因的發現功能基因表達提供了可靠依據[6]。但是當前尚未見到與羅氏沼蝦基因組序,蛋白質編碼密碼子的偏好性研究分析,近年來隨著深度測序技術的進一步發展,轉錄組測序為密碼子偏好性的進一步研究提供了數據來源,本研究以羅氏沼蝦卵巢組織轉錄組測序結果為研究數據,對羅氏沼蝦功能基因的密碼子偏好性特征進行分析,以期為羅氏沼蝦新基因測序功能基因組研究、外源基因密碼子的改造以及優化打下理論基礎,同時促進分子育種方面的深入研究。
1 ? ?材料與方法
1.1 ? ?試驗材料
以本課題組羅氏沼蝦卵巢組織的轉錄組結果為原始數據,為減少樣本誤差,篩選出6 629條具有完整開放閱讀框的序列,每條序列堿基長度均大于300 bp,并且以ATG為起始密碼子,以TAA、TAG或TGA為終止密碼子,且序列中間無N或者終止密碼子。
1.2 ? ?試驗方法
1.2.1 ? ?密碼子偏好性相關研究。通過在線CUSP程序以及Codon W 1.4.4(網址為http://codonw.sourceforge.net)對羅氏沼蝦6 629條具有完整閱讀框的序列進行密碼子組成和偏好性研究。計算GC、GC1、GC2、GC3、GC3s、氨基酸數(L_AA)、最優密碼子使用頻率(即FOP)、密碼子適應指數(即CAI)、密碼子偏愛指數(即CBI)、有效密碼子數(即ENC)、芳香性(aromo)、平均親水性值(gravy)等。
1.2.2 ? ?中性分析。中性分析(neutrality plots)是以GC12(GC1和GC2的平均值)為縱坐標、以GC3為橫坐標繪圖,分析密碼這3個點位上GC含量之間的聯系,從而研究密碼子在突變作用和選擇中心理論作出的貢獻[7]。若GC3與GC12兩者間具有內在的關聯,表示密碼子的使用偏好性在進化和遺傳過程中受到突變作用的影響;如果兩者不存在相關性,則表示在自然進化的有關過程中,自然選擇壓力在一定的程度上影響了該偏好性[8]。
1.2.3 ? ?ENC曲線分析。ENC曲線是以GC3s為橫坐標、以ENC為縱坐標,繪制ENC-GC3s分布圖,用來研究對該偏好性產生影響的主要因子,包括突變偏向性和突變壓力等[9]。一些密碼子僅受到突變偏向性即GC的約束,其ENC會在標準曲線的上方或附近位置。如果ENC在曲線的下方或遠離曲線的位置,那么說明自然選擇對該偏好性發揮了關鍵性作用。
1.2.4 ? ?最優密碼子分析。它的確定過程如下[10]:針對以大小來排序的ENC數據,進行的下限及上限相關區域均為10%的基因編碼區序列的有效選擇,然后分析及對比它們各自的RSCU值,分析確定最優密碼子。如果兩者RSCU值之差大于0.3,此外對于低表達以及高表達基因子集中,RSCU分別小于或是超過1.0,那么就認為它是最優密碼子[11]。
2 ? ?結果與分析
2.1 ? ?羅氏沼蝦轉錄組密碼子組成和使用
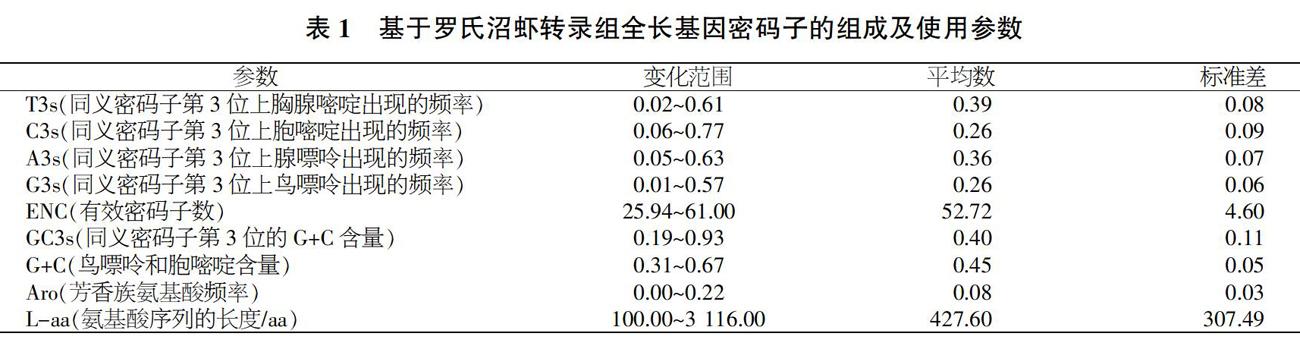
羅氏沼蝦轉錄組6 629個全長基因序列的長度變化范圍為303~9 351 bp,平均值為1 284 bp,所包含基因的G+C含量變化范圍為0.31~0.67,平均為0.45(表1);同義密碼子第3位G+C的含量變化范圍為0.19~0.93,平均數為0.40。這些數據證實,在其第三位核苷酸中,相應的GC含量并不存在非常顯著的偏好性;然而與GC總含量相比,各個基因之間所對應的GC3含量則有著更為廣闊的分布區間范圍。從密碼子第3位的堿基組成來看,第3位的堿基含量非平均分布,其分布趨勢是T含量最高,A含量次之,G和C含量較低;芳香族氨基酸(Aro)的頻率范圍為0.00~0.22。
羅氏沼蝦密碼子第1、2、3位的GC含量分別為51.58%、40.19%、41.24%,其第1位的GC含量高于第2、3位,這表明第1位密碼子是密碼子發生偏向性的主要作用力之一。
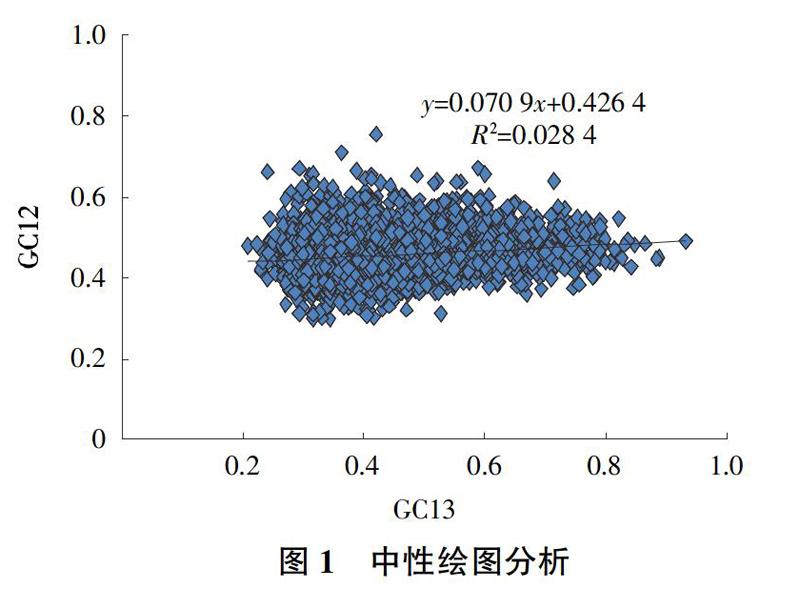
2.2 ? ?中性分析
中性分析(Neutrality analysis)可以有效分析GC12和GC13之間的相對聯系,從而可以更好地分析在密碼子偏好性使用過程中突變作用和選擇中性理論所做出的貢獻,中性分析可以有效分析密碼子的3個相對位置關系(GC12 vs GC3)(圖1)。分析結果表明,羅氏沼蝦基因組中的基因GC3含量的變化范圍(0.298 5~0.754 5)和GC12含量的變化范圍(0.207~0.933)都相對比較寬,并且GC3與GC12相關系數是0.168 5,相關性不顯著,同時回歸曲線斜率是0.070 9。
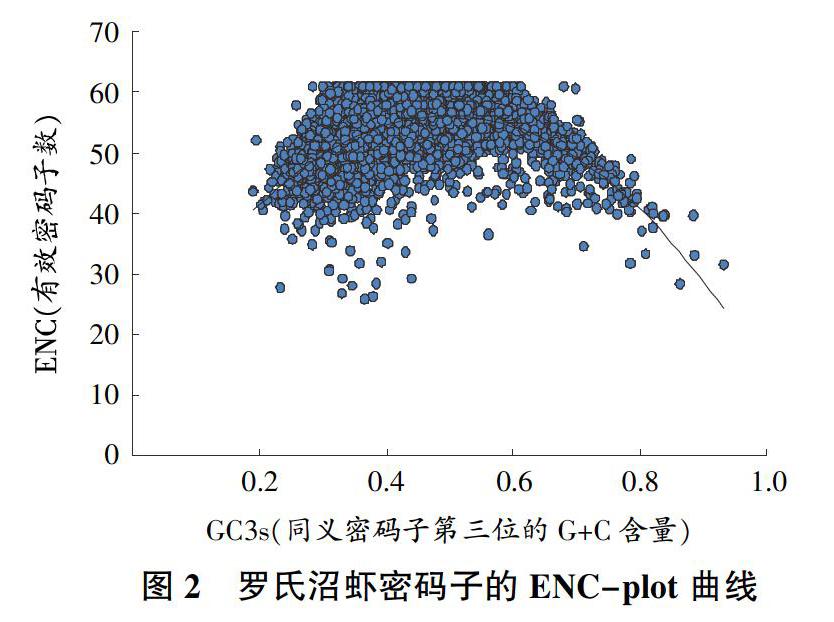
2.3 ? ?羅氏沼蝦密碼子使用參數相關性研究
以GC3s值為X軸、ENC值為Y軸,用SPSS 20.0繪制散點圖,所作出的連續性曲線反映了在沒有選擇壓力調節之下,兩者之間的相互關系,如圖2所示,大多數經位點分布偏離希望曲線。密碼子使用的影響因素包括自然選擇以及核苷酸組成方面的偏好等,如果同義密碼子偏向以C或G作為其結尾,且GC3含量較高,以及那些處在曲線下面部分的基因,它們的該類偏好性都較為明顯。因此,圖2中ENC比例偏低,而如果基因密碼子出于曲線的上方區域,則其有相對較低的偏好性,因而主要傾向以隨機的方式來進行密碼子的使用。
2.4 ? ?確定羅氏沼蝦的高頻密碼子
本文針對16 086個密碼子(來自篩選得到的6 629條表達基因),深入地分析了同義密碼子相對使用頻率(RFSC),發現對于各類密碼子而言,它們具有明顯不同的RFSC值(表2)。此處利用了高頻密碼子的相關篩選技術。經過分析,確定高頻密碼子為GAU、GAA、UUU、AAU、CCA,同義密碼子2個大于0.6、3個大于0.5、4個大于37.5、6個大于25。
2.5 ? ?密碼子使用頻率研究和最優密碼子的確定
相對頻率(RSCU)被認為是科學進行該類偏好性衡量的重要指標。如果RSCU>1,說明密碼子使用頻率較高;而如果RSCU<1,說明其使用頻率較低;假如RSCU=1,則意味著其使用不具備明顯的偏好性。
最優密碼子的確定參考STENICO等[12]的方法,最終確定了16個最優密碼子,編碼10個氨基酸,分別為Gly、Arg、Ser、Asp、Ala、Val、Ile、Thr、Pro、Leu等。對于Leu,它具有3個最優密碼子,即Ala、Pro、Ser,Thr為2個。16個最優密碼子中,除UUG外,剩下的各個密碼子結尾均為A/T,這就意味著其最優密碼子主要偏好結尾為A/T的形式(表2)。
2.6 ? ?羅氏沼蝦和相關的模式生物的密碼子偏好性對比研究
根據數據進行羅氏沼蝦相關密碼子統計分析,并且和密碼子相關用法專業數據庫(來自Kazusa DNA研究中心)共享的不同生物(包括人類、果蠅以及大腸桿菌等)數據展開對比性研究。對于不同物種之間具體對應的密碼子使用偏好性方面所存在差異性的有效衡量而言,密碼子的使用頻率比值是非常重要的一項科學指標。如果比值≤0.5或≥2.0,說明存在著較大差異的該類偏好性;而如果比值在0.5~2.0 之間,則意味著該類偏好性比較相似[12]。
根據表3所列出的研究結果可以發現,羅氏沼蝦和其他物種在該類密碼子偏好性上存在著一定的區別。羅氏沼蝦與人、斑馬魚和酵母的相關使用頻率分別有11、6、5種密碼子的比值≥2.0或≤0.5;和大腸桿菌以及果蠅等2類生物有比較明顯的差異性,分別有24、23種密碼子的比值≥2.0或≤0.5;而與中華絨螯蟹相比,具有最高的差異性,比值≥2.0或≤0.5有39個。
3 ? ?結論與討論
通過對羅氏沼蝦相關的同義密碼子所具有的偏好性進行深入地科學分析,發現其偏向于通過T或者是A來進行結尾,和幾類生物進行對比,羅氏沼蝦具有明顯不同的密碼子偏好性。與果蠅、中華絨螯蟹等生物具有顯著的差異性,而與酵母最為接近,但目前中華絨螯蟹公布的數據有限,可能密碼子偏好性有所偏差,有待數據進一步完善。
物種進化過程中,對同義密碼子的使用并不相似,物種會在特定的環境中,由于物種的適應性而進化出相應的特定密碼子,從而形成密碼子的使用偏好性,自然選擇效應以及突變性的壓力都對該類偏好性的產生具有較大影響[13]。其他多種因子如GC含量[14]尤其是與GC3s(密碼子第3位的G+C)含量、tRNA的豐富度[15]、密碼子位置、基因長度[16]、環境壓力、群體數目、突變偏好性[17]、基因表達水平[18]、編碼蛋白的機構與功能、翻譯效率[19]、mRNA二級結構等,都會影響密碼子使用偏好性。本研究發現,羅氏沼蝦基因組密碼子的使用偏好性受到了自然選擇壓力的影響。
密碼子的使用是了解基因遺傳和進化事件的有用工具。密碼子使用模式的分析對于理解基因組的遺傳和進化特征是重要的。采用密碼子偏好性的科學研究分析,一方面有助于給基因表達提供更為科學合理的表達系統,另一方面還可以給通過密碼子的改造來進行基因表達量的有效提升提供重要的基礎研究根據[20]。
4 ? ?參考文獻
[1] 陳雪峰,王春琳,顧志敏,等.羅氏沼蝦卵巢發育不同時期轉錄組分析[J].海洋與湖沼,2019,50(2):398-408.
[2] 馮超.基于楊梅RNA-Seq的密碼子偏好性與果實品質功能基因轉錄特性分析[D].杭州:浙江大學,2014.
[3] 段曉克.壯體長春鳊線粒體基因組全長測定及團頭魴密碼子偏好模型分析[D].武漢:華中農業大學,2015.
[4] 張欣,尤宏爭,李榮妮,等.金魚密碼子用法特點分析[J].華北農學報,2017,32(增刊1):30-36.
[5] 姚琳,江艷華,李風鈴,等.太平洋牡蠣(Crassostrea gigas)類A-1,2-巖藻糖基轉移酶的密碼子優化與原核表達[J].漁業科學進展,2016,37(1):74-79.
[6] 李喜蓮,楊元杰,李倩,等.螯蝦次目功能基因密碼子偏好性研究[J].浙江農業學報,2014,26(4):862-867.
[7] 徐偉榮,張寧波,王振平.歐洲葡萄堿性螺旋-環-螺旋(bHLH)基因家族密碼子使用偏性分析[J].農業生物技術學報,2014,22(4):440-448.
[8] SUEOKAN.Translation-coupled violation of parity rule 2 in human genes is not the cause of heterogeneity of the DNA G+C content of third codon position[J].Gene,1999,238(1):53-58.
[9] 張太奎,起國海,葉紅蓮,等.石榴轉錄組密碼子使用偏向性[J].園藝學報,2017,44(4):675-690.
[10] JIANG Y,DENG F,WANG H,et al.An extensive analysis on the global codon usage pattern of baculoviruses[J].Archives of Virology,2008,153(12):2273-2282.
[11] STENICO M,LLOYD A T,SHARP P M.Codon usage in Caenorhabditis elegans:delineation of translational selection and mutational biases[J].Nucleic Acids Research,1994,22(13):2437-2446.
[12] 李平,白云鳳,馮瑞云,等.籽粒莧蘋果酸酶(Nad-Me)基因密碼子偏好性分析[J].應用與環境生物學報,2011,17(1):12-17.
[13] 牛元,徐瓊,王崳德,等.大花香水月季葉綠體基因組密碼子使用偏性分析[J].西北林學院學報,2018,33(3):123-130.
[14] FAN S X,FEI H J,LIU S Q,et al.The features of synonymous codon bias and GC-content relationship in human genes[J].Progress in Biochemistry & Biophysics,2002,29(3):411-414.
[15] OLEJNICZAK M,UHLENBECK O C.tRNA residues that have coevolved with their anticodon to ensure uniform and accurate codon recognition[J].Biochimie,2006,88(8):943-950.
[16] 孫錚,馬亮,ROBERT M,等.Wolbachia Pipientis wMel基因組水平上的密碼子使用分析[J].中國科學,2009,39(10):948-953.
[17] XUHUA X.Mutation and selection on the anticodon of tRNA genes in vertebrate mitochondrial genomes[J].Gene,2005,345(1):13-20.
[18] 石秀凡,黃京飛.人類基因同義密碼子偏好的特征以及與基因GC含量的關系[J].生物化學與生物物理進展,2002,29(3):411-414.
[19] ZALUCKI Y M,POWER P M,JENNINGS M P. Selection for efficient translation initiation biases codon usage at second amino acid position in secretory proteins[J].Nucleic Acids Research,2007,35(17):5748-5754.
[20] 劉慶慧,黃倢,韓文君.Wssv 3個編碼蛋白的基因密碼子偏愛性分析[J].海洋水產研究,2005,26(4):1-7.
基金項目 ? 國家重點研發計劃“藍色糧倉科技創新”專項(2018YFD090
1303);浙江省農業(水產)新品種選育重大科技專項(2016C02055-2)。
作者簡介 ? 慎佩晶(1987-),女,浙江湖州人,碩士,工程師。研究方向:水產動物遺傳育種。
通信作者
收稿日期 ? 2020-04-01
