Tim-3調控巨噬細胞極化在類風濕性關節炎中的研究進展
2020-09-22 00:57:32鐘玉梅陳洋羅小超周海燕
天津醫藥 2020年9期
關鍵詞:信號
鐘玉梅,陳洋,羅小超,周海燕
類風濕性關節炎(rheumatoid arthritis,RA)是以滑膜炎癥為主要臨床表現的慢性、難治性、自身免疫性疾病,目前藥物治療的效果欠佳,且該病有較高的致殘率,嚴重影響了患者的生活質量[1]。T細胞免疫球蛋白黏蛋白分子-3(T cell immunoglobulin and mucin-domain- containing molecule-3,Tim-3)作為免疫系統共刺激抑制受體家族成員,是免疫調節的關鍵調控分子。在RA疾病進展中,Tim-3在局部免疫和全身免疫中均發揮作用[2-3]。同時,Tim-3 作為巨噬細胞極化信號通路的調控分子,其對巨噬細胞極化具有確切的影響[4-5]。本文就近年來Tim-3 調控巨噬細胞極化在RA中的研究進展進行綜述,以增強對Tim-3/巨噬細胞極化在RA 發病機制中的認識,也為RA的研究提供新的思路和治療靶點。
1 Tim-3的研究進展
Tim-3 是近年新發現的 T 細胞表面分子[6],是 T細胞免疫球蛋白黏蛋白(Tim)家族的成員,主要是由先天性和適應性免疫細胞分泌的一組Ⅰ型跨膜蛋白[7-9]。Tim-3廣泛表達于T細胞、巨噬細胞、自然殺傷細胞等細胞表面,并且與其配體結合發揮免疫調節功能。目前越來越多的研究表明,Tim-3與RA易感性密切相關,可促進多種細胞內信號傳導和免疫反應,調節T細胞分化,在對抗炎癥和自身免疫性疾病中起到了重要作用[10-12]。
1.1 Tim-3 及其配體的生物學特性 McIntire 等[13]在小鼠11 號染色體Tapr基因座位上發現了Tim 基因家族,其所在的染色體區與自身免疫性和變態反應性疾病發生相關。Tim-3分子結構包括3部分:胞外段、跨膜段、胞內段。胞外段由IgV區及含31%絲氨酸和蘇氨酸殘基的黏蛋白樣結構組成,含有4個N連鎖的糖基化位點和5 個O 連鎖的糖基化位點;胞內段含有6個酪氨酸殘基,其中1個酪氨酸殘基是酪氨酸磷酸化基序(RSENIY)中的一部分,而RSENIY與Tim-3下游信號傳導有關。人和鼠的Tim-3總氨基酸有63%的同源性,兩者胞內段有77%的氨基酸同源。
半乳糖凝集素-9(galectin-9,Gal-9)最早被認為是Tim-3的配體,它在體內分布非常廣泛,尤其是免疫細胞上[14];主要參與細胞聚集、黏附、分化、凋亡、調節腫瘤轉移和炎癥反應等[15]。Gal-9 含有碳水化合物識別區,通過與Tim-3的IgV結構域中的碳水化合物結構相結合進而發揮生物學功能。Tim-3/Gal-9 信號通路在腫瘤免疫[16]、抗感染免疫[17]和移植免疫[18]等方面發揮著重要的作用。
1.2 Tim-3 與免疫性疾病 Tim-3 作為負性免疫調節共刺激分子,一方面參與調控免疫平衡,維持對自身抗原的免疫耐受,確保完成免疫應答,避免自身免疫性疾病的發生;另一方面通過調控外周組織中免疫應答的持續性和強度,從而避免過度免疫造成的機體自我損傷[3]。研究發現,在輔助性T 細胞1(Th1)介導的實驗性自身免疫性腦脊髓炎中,阻斷Tim-3 可加速疾病的發作,導致小鼠病情惡化[6]。Liu等[19]發現,Tim-3的基因多態性可增加自身免疫性疾病的發病風險。劉葉等[20]發現,Tim-3 抑制能有效調節支氣管哮喘患者Th17 和調節性T 細胞(Treg)的平衡,減輕患者的氣道炎癥。在對免疫性血小板減少癥患者的研究中發現,Tim-3 的表達水平明顯升高,且與干擾素(IFN)-γ、血小板水平呈正相關[21]。Tim-3基因敲除后,T細胞分泌腫瘤壞死因子(TNF)-α、IFN-γ的水平顯著增高,體外殺傷胃癌AGS-EBV 細胞的能力也明顯增強[22]。另有研究發現,單核細胞/巨噬細胞表達的Tim-3 可能是胃癌進展的重要機制[23]。
2 巨噬細胞M1/M2 極化信號通路失衡與RA 的關系
巨噬細胞浸潤是RA 病理過程中的一個突出特征,而巨噬細胞又是固有免疫和適應性免疫的重要組成部分[24-26]。當受到不同因素刺激時,巨噬細胞可極化成為功能相反的2種表型:M1型(促炎型)和M2型(抗炎型)。M1型可導致促炎細胞因子炎癥介質產生,包括TNF-α、白細胞介素(IL)-1β、IL-6、IL-12 和單核細胞趨化蛋白(MCP)-1;而極化為M2 型時產生精氨酸酶1(Arg1)、IL-10、血管內皮生長因子(VEGF)等,這與免疫調節、抗炎、組織修復密切相關。在正常情況下,M1/M2 處于平衡狀態,M2 分化充足,抑制M1 的分化[27-30],即所謂“正氣存內,邪不可干”的狀態。但M1 與M2 在一定條件下可以相互轉化,并調節機體炎癥反應應答。研究發現,RA 中存在M1、M2表型極化的失衡[31-32]。在RA疾病發生發展過程中,多種因素會打破巨噬細胞M1/M2 型的動態平衡,引起數量和比例失衡,導致M1 型促炎巨噬細胞不斷增多,從而加劇炎癥反應[33],邪實日益嚴重。因此,有效干預巨噬細胞M1 型轉為M2 型,使M1/M2 恢復平衡狀態,有利于促進炎癥消退及組織修復,正氣足則邪自安。
3 Tim-3與RA
3.1 Tim-3 的免疫調控與RA 發病的關系 研究發現,RA 患者外周血及滑膜液中CD4+和CD8+T 細胞表面Tim-3 的表達均較健康人明顯升高,且Tim-3水平與病情的嚴重程度呈負相關[34]。RA 發病過程中,位于CD4+和CD8+T 細胞表面的Tim-3 起著負性調節免疫功能的作用,抑制T 細胞產生IFN-γ 等促炎因子,促進T 細胞生成IL-4 等抑炎因子。Tim-3作為免疫系統的關鍵調節者,是免疫炎癥的紐帶,也是抑制RA炎癥反應的重要共刺激因子,已成為治療RA的一個關鍵突破口。
3.2 Tim-3是調節巨噬細胞M1/M2極化信號通路的誘導器 Tim-3信號傳導對巨噬細胞炎癥因子表達和極化具有確切的影響,Tim-3高表達于M2型巨噬細胞,可促進巨噬細胞向M2 型極化并抑制M1 型極化,從而上調巨噬細胞中抗炎因子的表達并抑制促炎因子的表達;抑制Tim-3的表達可有效促進M1型巨噬細胞極化,抑制M2型巨噬細胞極化[35-36]。實際上Tim-3 對巨噬細胞極化的調控涉及2 部分:一種是通過接頭蛋白直接調控,Tim-3與其配體Gal-9結合增強Tim-3 與信號轉導及轉錄激活因子1(STAT1)之間的結合,再通過STAT1/miR-155/細胞因子信號傳導抑制因子1(SOCS1)信號軸發揮對巨噬細胞極化的調控;另一種是與Gal-9 結合發揮Tim-3/Gal-9 信號通路對T 細胞免疫發揮抑制或促進的調節作用,通過不同免疫反應產生的產物如IFN- γ 或 IL-4 誘 導 巨 噬 細 胞 向 M1 或 M2 型極化[37-38]。
此外,也有文獻報道Tim-3 信號可以促進β 干擾素TIR結構域銜接蛋白(TRIF)和干擾素調控因子3(IRF3)水平,調節巨噬細胞M1極化[39]。目前已證實在葡聚糖硫酸鈉(DSS)誘導的結腸炎小鼠模型中,Tim-3抑制了M1型巨噬細胞的極化和IRF3的磷酸化水平,IRF3 是調節巨噬細胞極化的Toll 樣受體4(TLR-4)下游轉錄因子[40],但目前對該通路的研究還不夠深入。
3.2.1 Tim-3 直接調控巨噬細胞極化 STAT1 是Tim-3 直接調控巨噬細胞極化的胞內關鍵接頭蛋白。研究發現,Tim-3 通過其細胞內尾部的殘基Y256 和 Y263 直 接 結 合 STAT1 并 抑 制 STAT1/miR-155/SOCS1 信號軸而極化巨噬細胞[37]。在該途徑中,Gal-9與Tim-3的結合增強了Tim-3與STAT1之間的結合,STAT1 在miR-155 的上游起作用并促進miR-155 轉 錄 ,SOCS1 參 與 Tim-3/STAT1/miR-155下游的信號級聯并介導巨噬細胞最后極化。Tim-3通過磷酸化殘基Y256和Y263與Janus激酶(JAK)競爭結合STAT1,然后抑制STAT1磷酸化;STAT1磷酸化受到抑制后使miR-155 表達降低,進而對SOCS1表達的抑制作用降低,隨后IL-10 和Arg-1 表達增加,最終導致巨噬細胞向M2表型的極化。
3.2.2 Tim-3 間接調控巨噬細胞極化 Tim-3 還可通過調節T細胞產生的IFN-γ或IL-4等產物誘導巨噬細胞向M1型或M2型極化。IFN-γ 是Th1細胞分泌的主要細胞因子之一,Tim-3/Gal-9信號通路可抑制Th1 細胞產生IFN-γ,而IFN-γ 是巨噬細胞M1 型極化的誘導因子,促使巨噬細胞M1型極化[41]。IL-4是Th2 細胞產生的主要細胞因子之一,Tim-3/Gal-9信號通路可促進Th2 細胞產生IL-4,IL-4 同時也是巨噬細胞M2 型極化的誘導因子,可誘導巨噬細胞M2 型極化。M1 的激活途徑由STAT1 介導。IFN-γ通過其受體 IFNR-1 和 IFNGR-2 募集 JAK1 和 JAK2銜接子,激活STAT1 通路和干擾素調節因子(IRF);IFN-γ 誘導的信號轉導的主要介質是STAT1、JAK2和IRF1[42-43]。M2的激活途徑主要由STAT6介導,首先IL-4 通過與其受體IL-4Rα 等結合,受體二聚化(Ⅱ型受體)后分別激活Janus 激酶JAK1/JAK3 或JAK1/JAK2,從而募集胰島素受體底物(IRS)家族主要成員形成復合物;磷酸化的IRS 激活生長因子受體結合蛋白2(Grb2)和磷脂酰肌醇3-激酶(PI3K),進一步募集STAT6,并使STAT6發生酪氨酸磷酸化;磷酸化的STAT6形成二聚體,入核后,啟動相關基因的轉錄,使巨噬細胞逐步向 M2 極化[38,44-46]。Tim-3通過IFN-γ/IL-4 調節巨噬細胞極化,最終產生作用不同的產物(圖1)。
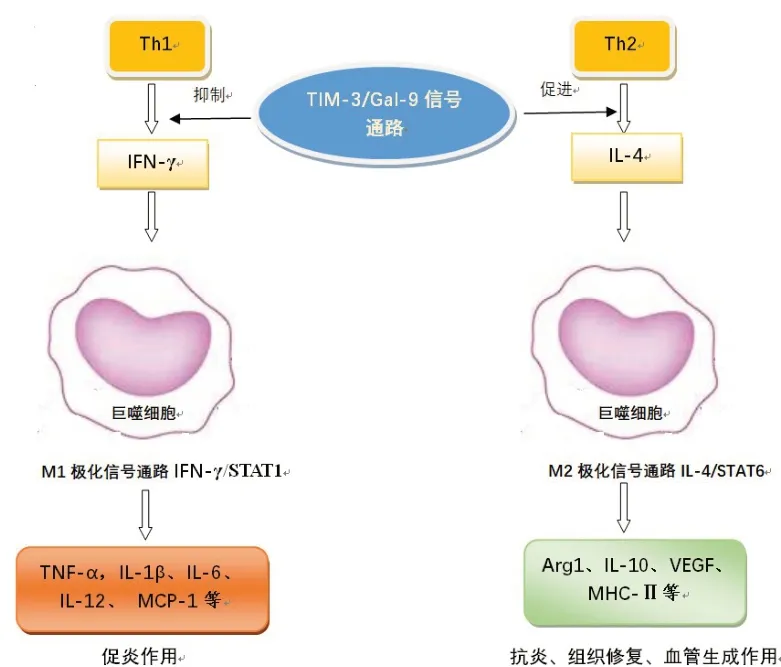
Fig.1 The flow chart of Tim-3 regulating the polarization of macrophage through IFN-γ/IL-4圖1 Tim-3通過IFN-γ/IL-4調節巨噬細胞極流程圖
4 小結
巨噬細胞作為固有免疫和適應性免疫的重要組成成分,其極化方向在RA病理過程中扮演了重要的角色。Tim-3 作為巨噬細胞極化信號通路的誘導器,其信號傳導可促進M2 型巨噬細胞極化并抑制M1 型極化,從而調節巨噬細胞中炎癥因子的表達,但其與RA 病理發展的相關性仍待進一步的研究。Tim-3 調控分子的研究不僅對深入揭示RA 的發病機制具有重要的指導作用,也為RA的臨床治療提供了新的靶點。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
