一磷酸鞘氨醇在哮喘炎癥細胞水平的研究進展①
2020-09-29 08:53:44劉函曄陳正愛劉衛東
中國免疫學雜志 2020年14期
關鍵詞:小鼠
劉函曄 陳正愛 劉衛東 崔 弘
(延邊大學基礎醫學院,延吉 133002)
哮喘是普遍的公共衛生問題,其發病率在世界范圍內不斷上升,特別是在工業化城市人口中,影響到全世界5%~10%的人口。然而,部分重癥哮喘患者對長效β-腎上腺素能受體激動劑產生依賴性或難以接受抗炎糖皮質激素等藥物,且處于嚴重發病和威脅生命的高風險中,僅不到50%的患者能達到可接受的臨床控制[1]。
1 S1P的來源
鞘磷脂是真核細胞脂質雙層中普遍存在的成分。鞘氨酰鞘氨醇(N-酰基鞘氨醇)是鞘脂的骨架,由神經酰胺合成和鞘脂周轉產生。鞘脂在分解代謝過程中,神經酰胺脫酰化產生鞘氨醇。鞘氨醇由鞘氨醇激酶1和2(SphK1和SphK 2)磷酸化形成一磷酸鞘氨醇(sphingosine 1-phosphate,S1P)[2]。S1P可以通過可逆去磷酸化和鞘氨醇化降解,也可以通過磷酸水解酶不可逆地裂解成磷酸乙醇胺和十六烷基化合物從而被S1P降解。
2 S1P及其受體的生物學功能
S1P含5種G蛋白偶聯受體,被稱為S1P1-5,大多數細胞表達一種或多種S1P受體亞型。S1P受體均可與 S1P以較高的親和力結合并通過受體間相互作用參與增殖、遷移和血管生成等關鍵的細胞過程[3]。S1P的不同受體均有不同的G蛋白家族的耦合特性:如S1P1有Gi/o家族耦合特性,S1P2有Gi/o,Gi2/i3以及GQ蛋白家族的特性,這種耦合特性可直調節小GTP酶(Rho、Rac和Ras)[4]。此外發現,內皮細胞中可檢測到豐富的S1P1,有研究發現成年小鼠主動脈內皮細胞質膜上的 S1P1與炎癥黏附分子豐度負相關,因此,S1P具有調節血管發展和微血管屏障功能[5]。在內皮細胞的促炎反應過程中,抑制 S1P及其受體可抑制過敏反應炎癥,S1P2受體的抑制完全消除了內皮細胞中TNF-α誘導的血管細胞黏附分子1(VCAM-1)和細胞間黏附分子1 (ICAM-1)表達。抑制S1P與S1P2結合可以直接抑制NF-kB通路參與細胞因子反應和由其誘導產生的內皮細胞促炎反應[6]。
3 S1P及其受體在哮喘細胞應答中作用
3.1S1P與肥大細胞 肥大細胞(mast cell,MC)是血液中的一種粒細胞,廣泛分布于皮膚和內臟黏膜下的微血管中,可分泌多種細胞因子。MCs長期以來被認為是哮喘過敏性氣道反應的關鍵效應細胞。接觸過敏原后,MCs結合IgE而活化,導致隨后的脫顆粒及生物活性介質釋放[7]。研究表明,S1P是MCs產生的鞘脂代謝物,且已被證實是過敏原誘導的MCs活化的重要調節劑,臨床研究表明S1P在慢性哮喘的發病機制中具有重要作用,其可破壞呼吸系統上皮細胞屏障,激活膽堿能受體[8]。在哮喘患者的支氣管肺泡灌洗液(BALF)中觀察到S1P水平升高,表明S1P在過敏反應中及MC依賴性炎癥反應中有重要作用。抗原刺激的 MCs將 S1P釋放到間質中可以顯著調節炎癥過程,且鞘氨醇激酶(SPHK)抑制劑治療顯著改善了小鼠哮喘模型的免疫反應,特定的SPHK抑制劑在過敏性哮喘的MC依賴性小鼠模型中可減弱氣道高反應性和炎癥[9]。microRNA(miRNA)是MCs發育的關鍵調節因子,在MC的生長、分化和遷移中起關鍵作用。研究表明,miR221通過IgE-抗原復合物調節細胞周期并刺激顆粒和細胞因子釋放MCs,且其在哮喘小鼠中明顯上調[10]。目前已經證明幾種miRNA可靶向影響S1P信號傳導途徑,如miR-130a-3p和miR-613靶標SphK2,miR133b和miR-363靶標S1PR1。截至筆者撰稿,未有實驗闡明miRNA、S1P、MCs間的關系,需要對其進行進一步研究。
3.2S1P與淋巴細胞 淋巴細胞中,Th2淋巴細胞驅動免疫炎癥反應并引發過敏性哮喘。轉錄因子 GATA3是 Th2淋巴細胞表達的關鍵轉錄因子,其參與 Th2細胞因子的合成,如 IL-4,IL-5,IL-9和 IL-13。這些細胞因子刺激與過敏級聯相關的其他免疫細胞的成熟,如嗜酸性粒細胞和MCs。其中IL-4和IL-13驅動免疫球蛋白類轉向IgE作用于B淋巴細胞[11]。在慢性過敏性哮喘中,IgE激活的MCs分泌嗜酸性粒細胞,如 IL-3,IL-4、IL-5、和粒細胞-巨噬細胞集落刺( GM-CSF1)等[12]。S1P是淋巴細胞從次級淋巴器官進入體循環的主要調節劑。血漿中的S1P主要由紅細胞和血管內皮細胞產生,淋巴樣S1P由淋巴管內皮細胞分泌。由于S1P裂解酶的高活性,組織中僅能維持較低水平的S1P。大多數血漿S1P可與高密度脂蛋白結合,通過調節淋巴細胞生成在免疫穩態中起重要作用,淋巴結中B細胞和T細胞及來自胸腺的成熟 T細胞和天然殺傷T細胞的排泄都需要S1P表達。S1P信號傳導在體內外均能刺激TH17分化。研究發現,在S1P1的C末端區域具有磷酸化缺陷的小鼠在實驗性自身免疫性腦脊髓炎(EAE)模型中顯示了TH17-顯性免疫應答和增強的神經炎癥。在該模型中,促炎細胞因子IL-6和IL-17及關鍵的TH17轉錄因子p-Stat3也得到了增強[13]。S1P1在癌癥免疫中可介導Stat3激活,而Stat3可直接促進S1P1表達[14]。
3.3S1P與其他細胞 除了上述免疫細胞外,多種細胞還可參與哮喘炎癥反應,如巨噬細胞,樹突狀細胞(DC),嗜堿性粒細胞等。研究表明巨噬細胞表達高水平的S1P,其激活S1P2和S1P3并觸發促炎介質CCL2,IL-1β和IL-18表達[15]。Arlt等[16]的研究表明,脂多糖(LPS)誘導成熟期DC暴露于S1P或FTY720可降低IL-12產生。S1P4可調節 DC分化,并可能影響 TH17細胞分化,在自身免疫中發揮潛在作用[17]。在肺動脈內皮細胞中S1P可以增加連環蛋白和概念蛋白含量,并增加細胞的跨細胞電阻,使其黏附連接結構穩定;也可直接影響巨噬細胞精氨酸和誘導型一氧化氮合酶活性。除了促進精氨酸合成外,TGF-β和 IL-10的促炎細胞因子也在S1P刺激的巨噬細胞中起作用。由此可見,S1P促進了一種新的巨噬細胞表型[18]。另有研究表明,與表征載脂蛋白M(APOM)結合的S1P在內皮細胞中發揮抗凋亡作用[19]。
4 S1P及其受體在哮喘氣道平滑肌及氣道重塑中的作用
S1P具有調節炎癥和氣道重塑期間氣道平滑肌功能,已經提出S1P通過刺激人ASM細胞增殖影響氣道重塑,其中一種機制是通過S1P通過RhoA依賴性抑制肌球蛋白磷酸酶[20]。調節氣道抵抗力的毒蕈堿受體(MR)信號傳導與哮喘相關MR下游信號傳導導致外周氣道收縮,該過程涉及SphK的激活和細胞內Ca2+水平的釋放。有研究表明S1P全身性給藥提高小鼠整個肺模型的氣道阻力和膽堿能活性。通過微陣列分析鑒定氣道平滑肌細胞中可經S1P調控的有相關文獻報道的88個基因的兩倍或更多,包括參與細胞增殖和氣道重塑的基因(HBEGF,TGFB3,TXNIP,PLAUR,SERPINE1)、細胞內信號傳導基因(RGS4,RGS2,DUSP5,MAP2K3,DGKH)和轉錄調控基因(NR4A1,NR4A3,EGR3,FOSB)[21]。已有研究表明HBEGF、RGS4和PLAUR等在哮喘患者氣道中上調。S1P的五種受體在氣道平滑肌中可表達三種(S1P1-3),S1P2和S1P3是平滑肌細胞中基因表達所必需的。但與S1P3相反,S1P2不通過鈣發出信號,但可能與G12/13偶聯并激活細胞中的Rho相關激酶途徑[21]。
5 S1P受體激動劑、拮抗劑
根據S1P及其受體在機體內產生的生物學反應,可通過拮抗或激動S1P相關受體來抑制哮喘或更為嚴重的氣道重塑等。FTY-720是S1P的合成類似物,主要激動S1P1、S1P3、S1P4、S1P5受體,經FTY-720治療后哮喘小鼠Th2相關轉錄因子受到抑制,進一步證明其可消除急性過敏原激發后的過敏性炎癥和氣道高反應性[22]。SEW-2871是一種S1P1選擇性激動劑,可參與多種免疫炎癥反應過程,SEW-2871處理的小鼠中TNF-α、IFN-γ水平顯著下降,提示其在淋巴細胞運輸和發育的調節中起關鍵作用[23]。JTE-013是一種S1P2拮抗劑,通過抑制S1P2介導的支氣管上皮NF-κB活化和CCL 3產生抑制哮喘變應性反應。除上述受體選擇性激動劑和拮抗劑外列出如SEW-2871(S1P1選擇性激動劑)、VPC-44116(S1P1拮抗劑)等受體激動劑及拮抗劑(表1)。
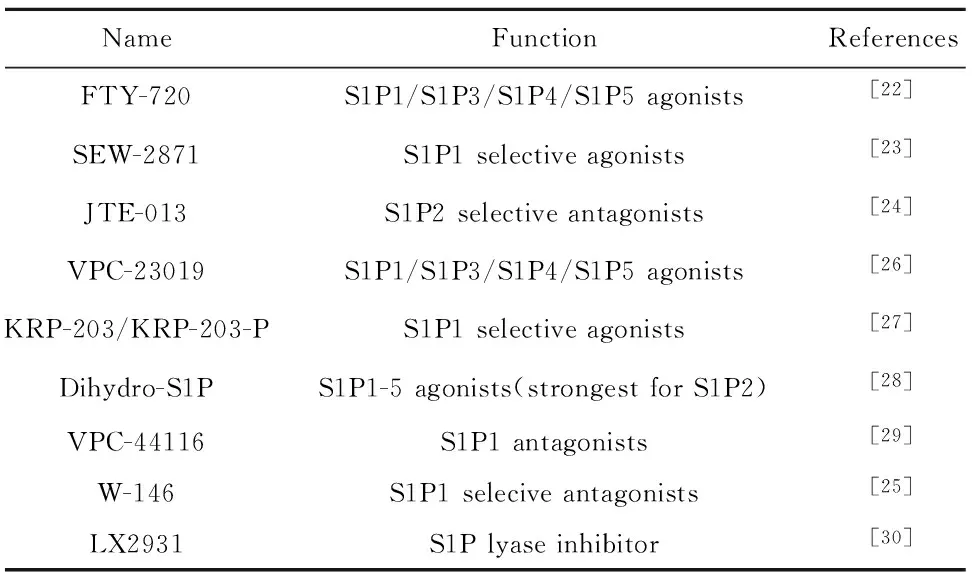
表1 S1P受體激動劑/拮抗劑
6 展望
綜上所述,S1P可增加炎癥產生并破壞呼吸系統上皮細胞屏障完整性,在細胞內外均影響炎癥發生,另一方面S1P可直接參與平滑肌的生物學反應, 影響氣道重塑。反復接觸變應原可誘發支氣管哮喘急性發作、炎癥因子分泌及氣道重塑,成為哮喘發生的主要特征,且氣道重塑是哮喘慢性化、持續化、嚴重化的病理基礎。激活的膽堿能受體等也與哮喘和慢性阻塞性肺病有關。本綜述對S1P及其受體在哮喘中作用進行歸納整理并推測對其受體及相關下游炎癥通路的研究是切實可行的,但仍有部分S1P受體激動劑和拮抗劑與哮喘關系尚未深入研究,可通過對其S1P/S1PR等下游通路的進一步檢測和對相關炎癥因子的表達推測其對炎癥的抑制作用。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
