不同實生變異和常規檵木光合特性的比較
2020-09-30 02:51:56衛東炎林
經濟林研究 2020年3期
,, , , ,衛東,,炎林
(1.湖南農業大學 a.園藝學院;b.湖南省中亞熱帶優質花木繁育與利用工程技術研究中心;c.教育學院,湖南 長沙 410128;2.園林花卉種質資源與綜合利用湖南省重點實驗室,湖南 長沙 410125)
檵木Loropetalum chinense為金縷梅科檵木屬常綠灌木或小喬木,目前在園林綠化工程和城鎮化建設工程中得到廣泛栽植和應用[1]。變異植物滿足了人們對觀賞植物新、特、奇的要求,且目前地被植物在園林綠化中得到越來越廣泛的應用,具備特異性狀的檵木屬變異植株將更受市場歡迎。近年來,關于檵木屬植物的研究主要集中在遺傳多樣性、生理特性、脅迫生理、繁殖育種和開發利用等方面[2-5]。
光合作用是植物進行生長發育、開花結果的生理生化基礎,其影響因子有光照、環境CO2濃度、環境溫度、光受體、礦質營養等[6-7]。近年來,通過改變植物中光合相關過程來提高生物量的方法受到了極大的關注。植物為了更好地適應其所處的環境條件,可根據相應的環境條件來采取適應策略,包括光合作用和新陳代謝的變化[8]。朱運欽等[9]在對不同株型桃樹光合特性的研究中發現,在普通型、半矮化型和垂枝型桃品種間光補償點的差異較顯著;緊湊型桃的光飽和點最高,垂枝型桃次之,普通型桃最低。姜衛兵等[10]經研究發現,與一般蘋果樹相比,短枝型蘋果樹具備較高的光飽和點和較低的光補償點。張光弟等[11]在矮化型觀賞植物蘋果矮化砧B9 和N29 光合特性的研究中發現,N29 的弱光利用能力高于B9,而B9 的高光適應能力較強。也有關于檵木屬植物光合特性的研究報道。陳曉林等[12]和王小青等[13]進行了不同種類紅花檵木光合特性的比較研究,結果表明不同品種的光合特性和適應環境的能力不同;董佳麗等[14]比較了紅花檵木實生后代光合特性的差異,認為后代與其母本的光合特性差異顯著;謝智華等[15]探究了不同酸度營養液對紅花檵木光合特性的影響,認為春季紅花檵木新枝在pH5.5 的弱酸性水培環境中光合表現最佳;桂克印等[16]經研究認為,外界物質對紅花檵木葉片光合特性的影響較小;化香平等[17]探究了南京地區紅花檵木與紅葉石楠光合特性的差異,結果表明紅花檵木的葉綠素參數均比紅葉石楠高,但其光合能力弱于紅葉石楠。
檵木屬植物作為具有多種園林用途的彩葉樹種,其株型與光合特性之間是否存在直接關系,及其光合特性是否影響其生態習性等仍有待探明。而關于檵木屬變異植物光合特性方面的研究鮮見報道。探討檵木屬植物匍匐和垂枝變異類型在光合特性和葉綠素熒光特性方面的改變,對檵木屬植物的品種選育具有較大的理論和實踐意義。因此,有必要加強檵木屬變異植株光合特性分子調節機制方面的研究。本試驗中通過對檵木屬不同變異株系光響應曲線的測定,結合葉片葉綠素熒光等相關光合生理數據,研究其在同等環境條件下的光合特性差異,分析檵木屬不同實生變異植株的光合能力,以期探明檵木屬植物株型與光合特性之間的關系,為改良檵木屬植物的栽培條件提供參考依據。
1 材料與方法
1.1 試驗地概況
試驗地設在湖南省長沙市湖南農業大學花卉基地(113°08′E,28°18′N)。長沙市坐落在湖南省東部偏北的湘江下游和長沙盆地西緣地區,屬亞熱帶季風氣候。年平均氣溫17.2 ℃,年均降水量1 361.6 mm,年平均日照1 200 ~1 600 h,無霜期年平均約275 d,霜期一般出現在11月至次年3月。氣候溫和,降水充裕,雨熱同期,四季明顯[18]。
1.2 試驗材料
試驗材料來源于湖南農業大學花卉基地紅花檵木種質資源圃中自然授粉的‘花葉檵木2號’的實生苗,通過觀察,從實生苗中篩選出2 個枝形有明顯外觀變異的優良單株‘臥龍’和‘42號’,分別編號為Y1 和Y2,以‘瀏陽紅’(Y3)和‘大葉紅’(DYH)為對照(圖1)。變異材料Y1 的特征為:株型矮小,分枝密,枝條蜷曲呈匍匐生長;葉較小,橢圓形,頂端尖,葉面有光澤且星狀毛較少;常綠不落葉,葉色為深淺不一的綠色,新葉帶有老葉綠色的條帶(色暈、色塊);花為白色。變異材料Y2 的特征為:枝條細軟下垂,似垂柳;葉較小,長卵形,新葉為墨綠色,老葉秋季變紅掉落;花為白色。10月中旬至11月中旬,選取生長狀況良好且無病蟲害的向陽植株的葉片作為研究對象,每個處理重復3 次。

圖1 變異檵木與常規檵木試材的形態Fig.1 Morphological characteristics of variations and normal materials of L.chinense
1.3 試驗方法
1.3.1 光響應特征參數的測定和光合-光響應曲線的繪制
09:00—16:00,每種植物隨機選擇3 片葉,使用LICOR 公司(美國)生產的LI-6400 便攜式光合儀測定光響應曲線。測定前對葉片進行光誘導10 min,待其完成光誘導并達到穩態后開始測定。測定時大氣溫度為20 ~25 ℃,以大氣作為CO2源,制作空氣緩沖瓶,來保持CO2濃度的相對穩定,流速為500 μmol/s,內置光照強度(PAR)梯度設置為0、20、50、80、100、200、500、800、1 000、1 200、1 500、1 800 μmol/(m2·s)。使用Photosyn Assistant 軟件對數據進行處理,分析植株的光補償點、光飽和點、初始表觀量子效率和最大凈光合速率(Pnmax)等數據,根據測定的凈光合速率(Pn),繪制光合-光響應曲線。
1.3.2 葉綠素熒光參數的測定和葉綠素熒光誘導動力學曲線的繪制
2019年10月25日,使用Fluor Pen FP110 手持式葉綠素熒光儀進行葉綠素熒光參數的活體測定。分別選取Y1、Y2、DYH、Y3 同一部位受光一致且為從形態學上端數第4 片健康功能葉進行測定,每株選3 片葉,取其平均值。每次測定前,將待測葉片擦拭干凈,使用葉片夾夾住且不夾在葉片的主葉脈上,進行20 min 暗處理。暗處理結束后開始測定葉綠素熒光參數,測量過程中使儀器探頭、葉片夾和葉片三者緊密貼合。測定的葉綠素熒光參數主要包括初始熒光(Fo)、可變熒光(Fv)和最大熒光(Fm)。計算PS Ⅱ的潛在活性(Fv/Fo)、PS Ⅱ最大光化學效率(Fv/Fm)、非光化學淬滅(NPQ)和光化學淬滅(PQ)。根據葉綠素熒光參數,參照付春霞等[19]和楊宋琪等[20]的方法,繪制葉綠素熒光誘導動力學曲線。
1.4 數據處理
用Excel 2010 軟件整理數據,用SPSS 22.0軟件進行單因素方差分析,用LSD 法和Duncan法對各參數進行差異顯著性檢驗和多重比較,用Origin 8.0 軟件作圖。
2 結果與分析
2.1 實生變異和常規檵木植株光響應特征參數的比較
選用直角雙曲線修正模型[21]描述實生變異及常規檵木植株的光合-光響應曲線,得出4 種植株的光響應特征參數(表1)。由表1可知,Y3、DYH 和Y2 的Pnmax分別為9.655、9.142 和6.704 μmol/(m2·s),差異不顯著(P>0.05),而Y1 的Pnmax為15.864 μmol/(m2·s),是前者的1.64 ~2.37 倍,差異顯著(P<0.05),表明Y1的光合能力較強。在4 種植株中,Y1 的初始表觀量子效率達到0.066,表明其捕獲光子能力較強,其他3 種植株間無顯著差異,且均與Y1 差異顯著(P<0.05)。4 種植株的光飽和點不具有顯著差異,表明其對強光的適應能力基本相同。Y2 的光補償點和暗呼吸速率與其他3 種植株差異顯著,表明其對弱光的利用能力較強,在無光條件下具備較小的呼吸速率。
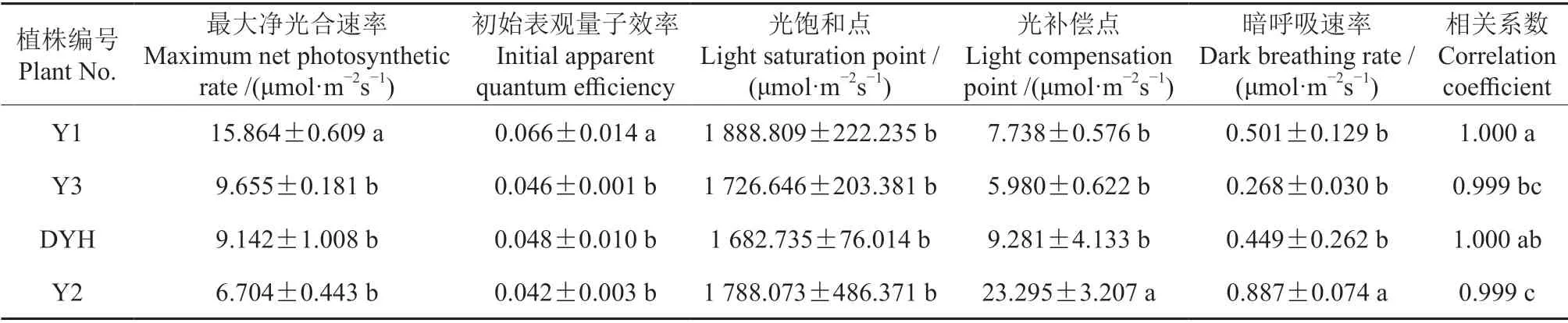
表1 實生變異和常規檵木植株的光響應特征參數(直角雙曲線修正模型)?Table1 Comparison of light response characteristic parameters of seed propagated variations and normal plants of L.chinense (modified rectangular hyperbolic model)
2.2 實生變異和常規檵木植株光合-光響應曲線的比較
光合-光響應曲線反映了植物的凈光合速率隨光照強度增減而變化的規律,實生變異及正常檵木植株的光合-光響應曲線如圖2所示。由圖2可見,4 種檵木植株的光合-光響應曲線的變化趨勢基本一致,Pn隨PAR 的增強而逐漸增加。整體上,各植株光合速率由高到低依次為Y1、Y3、DYH、Y2。
2.3 實生變異和常規檵木植株葉綠素熒光參數的比較
葉綠素熒光是一種非損傷探針,用來研究植物的光合作用非常有效[22-23]。實生變異及常規檵木植株的葉綠素熒光參數見表2。由表2可知,Y1的Fo值最大,為8 604.67,Y1 與Y2 的Fo值無顯著差異(P>0.05),但與Y3 和DYH 差異顯著(P<0.05)。Y1 的Fm值和Fv值最大,分別為45 417.67 和36 813.00,與其他植株間差異顯著(P<0.05),其他植株間差異均不顯著(P>0.05);在Y1、Y3 和DYH 三者間,Fv/Fm值、Fv/Fo值和Fm/Fo值無顯著差異(P>0.05),但三者的上述指標均與Y2 存在顯著差異(P<0.05)。
PS Ⅱ所吸收的光能無法被光合作用消耗,將多余光能用熱的形式耗散,即NPQ,其反映了植物的光保護能力[24-26]。光照下,實生變異和常規檵木植株NPQ 和PQ 的變化過程如圖3所示。由圖3可見,隨著光照時間延長,Y1 和DYH 的NPQ 一直呈遞增趨勢,表明其熱耗散能力較強,葉綠素吸收的光能向光合電子傳遞消耗較多,光合能力較強,更有利于抵制光抑制對光合機構的損害。Y3 在光照的40 s 內有快速上升的過程,然后一直保持平緩上升,至光照160 s 時呈下降趨勢。Y2 的NPQ 在40 s 內增長后出現了“降低—升高—降低—升高—降低”的趨勢。PQ 表現了PS Ⅱ反應中心開放部分的占比,反映植物在一定光環境下應用光能來進行光化學轉化的能力,代表了其光合效率[27]。Y1 的PQ 值整體上呈現緩慢上升的趨勢,而Y2、Y3 和DYH 整體上呈現先升高、再降低的趨勢,其中Y2 的PQ 值較高,表明其PS Ⅱ反應中心電子傳遞活性較大,而且PS Ⅱ反應中心開放的比例較大。
2.4 實生變異和常規檵木植株葉綠素熒光誘導動力學曲線的比較
植物葉綠素熒光誘導動力學曲線可反映PS Ⅱ反應中心原初光化學反應的相關信息。J 期反映電子從QA-到QB的積累,I期代表電子從QA到QB-的積累,P 期被認為是電子從QA-到QB2-的積累[28]。實生變異及常規檵木植株的葉綠素熒光誘導動力學曲線如圖4所示。由圖4可見,各植株葉片的葉綠素熒光誘導動力學曲線各時間點和形態有一定差別。O 點和P 點為Y1 的熒光強度最高,說明其光抑制程度較小,Y1 與其他3 種植株間存在差異,說明株型的不同可造成檵木屬植物葉綠素熒光強度的差異。自O 點差異開始,隨著J 點和I點差異的積累,到達P 點時差異達到最大。在對葉片進行快速熒光動力學誘導時,光照面積和強度不同,可能會使得葉綠素熒光誘導動力學曲線出現較大差別。在I—P 段,Y2 與DYH 的熒光強度并未有太大差異,說明其較多的光能用于轉變成化學能。
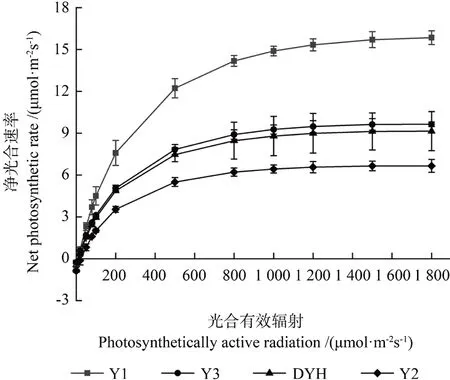
圖2 實生變異和常規檵木植株的光合-光響應曲線Fig.2 Comparison of photosynthetic light response curve of seed propagated variations and normal plants of L.chinense
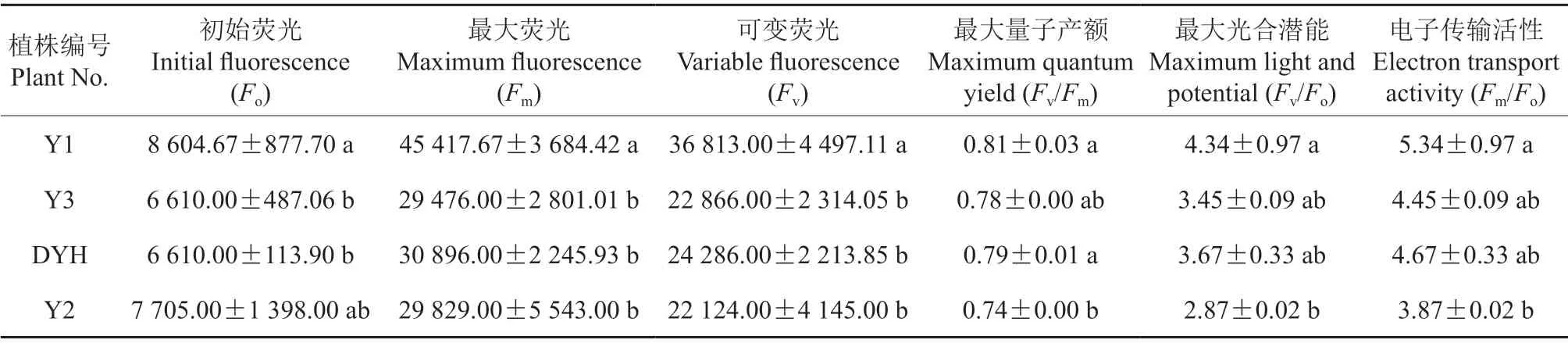
表2 實生變異和常規檵木植株的葉綠素熒光參數Table2 Comparison of chlorophyll fluorescence parameters of seed propagated variations and normal plants of L.chinense
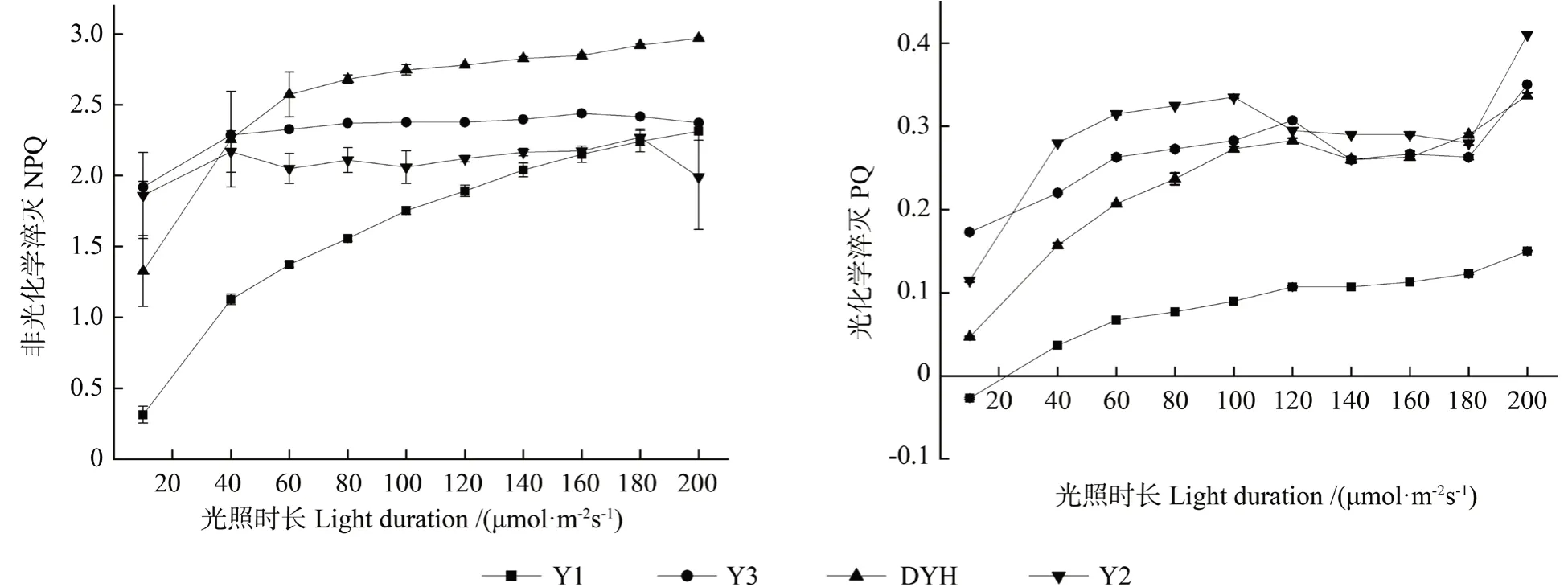
圖3 實生變異和常規檵木植株NPQ 與PQ 的變化曲線Fig.3 NPQ and QP variation curve of seed propagated variations and normal plants of L.chinense
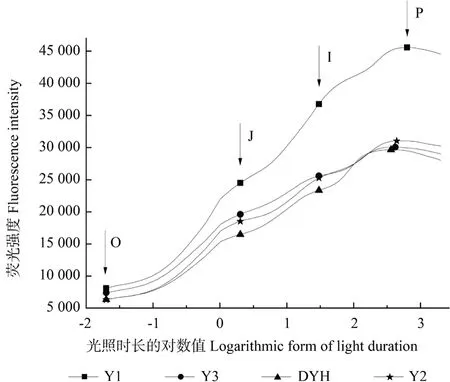
圖4 實生變異和常規檵木植株的葉綠素熒光誘導動力學曲線Fig.4 JIP test curves of seed propagated variations and normal plants of L.chinense
3 結論與討論
檵木屬植物實生變異會影響其光合效率和生態適應性,從而導致光合能力的較大改變。通過對4 種檵木植株葉綠素熒光參數和光響應參數的比較,發現匍匐突變植株‘臥龍’的最大凈光合速率、光飽和點、初始表觀量子效率均最大,光補償點較低,電子傳遞效率最大,光合性能較好,熱耗散較強,在強光下受到光抑制作用小,也能較好地利用弱光,而垂枝突變植株‘42號’的光合適應能力較對照植株稍弱。本試驗中,實生變異類型‘臥龍’的光合性能最強,能通過光合作用充分利用光能,積累大量有機物質,表現出對環境較強的適應性,其在園林景觀應用中的前景較好。在栽培時,因‘42號’較不耐陰,應首選在向陽坡適當種植,在進行城市美化景觀配植時應適度疏植。
光合作用作為影響植物生長發育的重要因素之一,受到外部環境條件和自身因素的綜合影響,其光合生理參數是科學制訂栽培措施的重要依據[29-30]。植物的光補償點和光飽和點是評估植物對光能的利用能力的可信指標,光飽和點越高且光補償點越低,表示其對光的適應范圍較大,光適應性較強[31-32]。本研究中,‘臥龍’和對照植株的光補償點較低,表明三者耐陰性較強,而‘42號’的光補償點較高,表明其最不耐陰。其中,光補償點參數也表明匍匐突變植株較對照更加耐陰,而垂枝突變植株較對照耐陰性更弱。一般認為,光飽和點高的植物能更加有效地利用強光,而‘臥龍’的光飽和點和最大凈光合速率均顯著高于其他3 種植株。因此‘臥龍’對光環境的適應性更廣,既能在強光下較好地利用光能,又對弱光具有一定的耐受性。崔永寧等[33]對大花黃牡丹變異材料光合特性的研究結果表明,變異品種的光補償點和最大凈光合速率均顯著高于同品種的非變異植株,這與本研究結論相一致。與對照植株相比較,匍匐突變植株的優勢更加突出,其光合能力和獨特株型更能滿足市場需求;垂枝突變植株的暗呼吸速率較對照植株更高,其光合能力與對照相差無幾,但消耗較多。
葉綠素熒光無損檢測技術被廣泛用于研究植物的光合生理和逆境脅迫生理,葉綠素熒光參數指標能夠反映植物葉片光系統Ⅱ對光能的吸收、傳遞和耗散等狀況[34-35]。Fo作為暗反應下的初始熒光,表示植物葉片的光合中心光系統Ⅱ在完全開放狀態下的熒光產量,與植物葉片的葉綠素濃度有關[36]。本研究中,‘臥龍’的Fo值最高,其次是‘42號’,說明二者的葉綠素含量比對照高,Fo值的上升能有效避免PS Ⅱ活性中心發生不可逆轉的毀壞,表明‘臥龍’和‘42號’的光合性能較好。Fm表示經過PS Ⅱ的電子傳遞情況,Fv表示PS Ⅱ的電子傳遞效率[37]。‘臥龍’的Fm值和Fv值在4 種植株中均是最高的,另外‘臥龍’的PQ 值不停上升卻低于其他3 種植株,說明其PS Ⅱ反應中心活性相對較大,電子傳遞效率較高,在強光環境下可較好地開啟熱耗散機制,讓植物免受光抑制的損傷。‘42號’的PQ 值較高,表明其有較高的光能利用效率。
根據葉綠素熒光誘導動力學曲線能夠分析出PS Ⅱ反應中心的活性,各植株的Fm值由高到低依次是‘臥龍’‘大葉紅’‘42號’‘瀏陽紅’,各植株的Fv值由高到低依次是‘臥龍’‘大葉紅’‘瀏陽紅’‘42號’。Fv/Fm表示植物潛在的最大光合能力,Fv/Fo代表光系統PS Ⅱ的潛在光化學活性,其值越高說明植物的光合性能越強,二者也是植物發生光抑制的敏感指標,可用于評估植物對環境的適應性,檢驗植物生長是否受到抑制[38-40]。一般植物不受脅迫的Fv/Fm值為0.75 ~0.85。本試驗中,‘42號’的Fv/Fm值小于0.75,說明此時葉片已經受到脅迫。‘42號’的NPQ 在光照120 ~180 s 時出現上升趨勢,是為了維護PS Ⅱ反應中心,而促發本身的保護機制來耗散掉多余的能量,而PQ 出現下降趨勢在一定程度上表示PS Ⅱ反應中心開放程度的下降,適應光合作用能力的下降[41],表明外界脅迫在一定程度上抑制了‘42號’的光合作用。‘臥龍’和對照的Fv/Fo值和Fm/Fo值明顯高于‘42號’,說明三者較‘42號’具有更高的PSⅡ潛在光化學活性和電子傳輸活性。有研究報道,逆境脅迫會使植物葉綠素熒光參數Fm、Fv/Fm、Fv/Fo和Fm/Fo降低,Fo升高[42-43],這與本試驗中‘42號’葉綠素熒光參數的變化相一致。與對照植株相比,匍匐突變植株‘臥龍’的整個光系統機制更強,能夠更好地適應環境;而垂枝突變植株‘42號’因秋季變色落葉形成自我保護機制,不利于植株整體光合作用的進行,導致與對照植株存在顯著差異。
由于受到本試驗中所選材料數量的限制,其生長年限不一致,僅設計了3 個重復處理,受植物生長階段等因素的影響,研究結果具有一定局限性。為了選育綜合性狀優良的檵木資源,后續將進一步研究檵木生長指標與光合特性的相關性。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
