鼎湖山錐栗木質(zhì)部形成及其對(duì)氣候的響應(yīng)
2020-10-09 14:31:50王婕余碧云黃建國(guó)
熱帶亞熱帶植物學(xué)報(bào) 2020年5期
王婕, 余碧云, 黃建國(guó)
(1. 中國(guó)科學(xué)院華南植物園退化生態(tài)系統(tǒng)植被恢復(fù)與管理重點(diǎn)實(shí)驗(yàn)室, 廣東省應(yīng)用植物學(xué)重點(diǎn)實(shí)驗(yàn)室, 廣州 510650; 2. 中國(guó)科學(xué)院核心植物園植物生態(tài)學(xué)科, 廣州 510650; 3. 中國(guó)科學(xué)院大學(xué), 北京 100049)
森林生態(tài)系統(tǒng)在全球碳循環(huán)中起著關(guān)鍵作用,被認(rèn)為是有助于緩解氣候變化的重要碳匯[1]。近年來(lái)溫度的持續(xù)上升和干旱的加劇,導(dǎo)致了森林的大面積死亡[2]。因此,弄清氣候變暖對(duì)森林生態(tài)系統(tǒng)的影響,對(duì)預(yù)測(cè)氣候變暖背景下森林生態(tài)系統(tǒng)可能發(fā)生的變化具有重要意義。樹(shù)木作為森林生態(tài)系統(tǒng)的重要組成部分,其生長(zhǎng)過(guò)程是一個(gè)重要的固碳過(guò)程,能直接影響森林群落組成和演替變化,進(jìn)而影響森林生態(tài)系統(tǒng)的發(fā)展和變化[3]。木質(zhì)部形成是樹(shù)木長(zhǎng)期碳匯的重要過(guò)程且易受到環(huán)境因素的影響[4]。通過(guò)對(duì)西伯利亞北部針葉樹(shù)的年際木質(zhì)部形成及其與氣候因子關(guān)系的研究,表明春季早期升溫與年際木質(zhì)部形成呈正相關(guān)[5]。而另一研究表明,在季節(jié)性干旱的熱帶森林中,水分是木質(zhì)部形成的主要驅(qū)動(dòng)因素[6]。因此,在不斷變化的環(huán)境條件下,木質(zhì)部的形成與氣候間的相互作用對(duì)評(píng)估氣候變化下森林的生產(chǎn)力和碳儲(chǔ)量具有重要意義。
樹(shù)木木質(zhì)部生長(zhǎng)的監(jiān)測(cè)對(duì)于了解森林生產(chǎn)力和氣候變化的關(guān)系具有十分重要的作用[7]。近年來(lái),相比于其他采樣方法,微樹(shù)芯采樣技術(shù)對(duì)樹(shù)木破壞性小、可連續(xù)采樣,并能觀察到高質(zhì)清晰的樣品,因此已被廣泛應(yīng)用于樹(shù)木木質(zhì)部生長(zhǎng)監(jiān)測(cè)的研究。Rossi等[8]利用微樹(shù)芯采樣技術(shù)建立了能預(yù)測(cè)全球變暖背景下木質(zhì)部活動(dòng)開(kāi)始、結(jié)束的時(shí)間以及持續(xù)變化時(shí)間的模型;Huang等[9]利用微樹(shù)芯采樣技術(shù)首次建立了定量樹(shù)木初級(jí)生長(zhǎng)與次級(jí)生長(zhǎng)關(guān)系的混合效應(yīng)模型,為更準(zhǔn)確預(yù)測(cè)森林生態(tài)系統(tǒng)生產(chǎn)力和碳平衡提供科學(xué)與理論依據(jù)。可見(jiàn),微樹(shù)芯采樣技術(shù)是研究樹(shù)木木質(zhì)部形成及其影響因素的有效方法。
北半球中高緯度地區(qū)的森林是一個(gè)巨大的碳匯,有助于延緩全球變暖的進(jìn)程。因此,前人的研究多集中于北半球的中高緯度高海拔地區(qū),亞熱帶地區(qū)樹(shù)木生長(zhǎng)的研究較少涉及。溫帶和北寒帶地區(qū)的氣候具有明顯的季節(jié)性差異,樹(shù)木生長(zhǎng)也表現(xiàn)出明顯的生長(zhǎng)季和非生長(zhǎng)季。Gricar等[10]加熱挪威云杉(Picea excels)的木質(zhì)部,結(jié)果表明溫度只在生長(zhǎng)季開(kāi)始時(shí)對(duì)樹(shù)木生長(zhǎng)有促進(jìn)作用,而在生長(zhǎng)季后期則無(wú)顯著作用, 說(shuō)明樹(shù)木生長(zhǎng)在生長(zhǎng)季前期和生長(zhǎng)季后期存在一定差異。中國(guó)南部亞熱帶地區(qū)森林生態(tài)系統(tǒng)的平均凈生產(chǎn)力(NEP)占全球的8%,面積占全球的近6%[11],是全球重要的碳匯[12]。目前,中國(guó)南部亞熱帶地區(qū)的森林樹(shù)木的木質(zhì)部形成及其對(duì)氣候變化的響應(yīng)尚不明確, 解決這一問(wèn)題對(duì)預(yù)測(cè)未來(lái)氣候變化背景下該地區(qū)森林生態(tài)系統(tǒng)的動(dòng)態(tài)過(guò)程具有重要意義。
闊葉林尤其是常綠闊葉林是亞熱帶地區(qū)的地帶性植被,常綠闊葉樹(shù)種在亞熱帶森林演替后期基本占主導(dǎo)地位[13]。闊葉樹(shù)地下根系復(fù)雜,更能充分吸收水分,涵養(yǎng)水源。常綠闊葉林樹(shù)種豐富,生態(tài)系統(tǒng)的復(fù)雜和多樣性孕育著豐富的物種多樣性和基因多樣性,因此,無(wú)論是在蓄水固土、物種保存還是環(huán)境保護(hù)方面闊葉樹(shù)都比針葉樹(shù)更有優(yōu)勢(shì)[14]。錐栗(Castanea henryi)是殼斗科(Fagaceae)栗屬植物,鼎湖山針闊葉混交林的主要建群種和優(yōu)勢(shì)樹(shù)種之一,也是我國(guó)特有的果材兩用樹(shù)種,其本身具有十分重要的生態(tài)和經(jīng)濟(jì)價(jià)值[15]。亞熱帶由于氣候溫和,樹(shù)木(尤其是闊葉樹(shù))連續(xù)生長(zhǎng)導(dǎo)致年輪界限不清晰,傳統(tǒng)的樹(shù)木年輪學(xué)方法難以對(duì)其進(jìn)行準(zhǔn)確的定年,本研究利用微樹(shù)芯采樣技術(shù),從微觀角度嘗試模擬闊葉樹(shù)錐栗木質(zhì)部在2015年的年內(nèi)生長(zhǎng)動(dòng)態(tài),并分析其對(duì)氣候的響應(yīng),為該地區(qū)森林的長(zhǎng)期監(jiān)測(cè)提供基礎(chǔ)資料,也為自然保護(hù)區(qū)森林的保護(hù)與管理提供理論依據(jù)。
1 研究區(qū)概況
研究區(qū)位于廣東省鼎湖山國(guó)家級(jí)自然保護(hù)區(qū)(112°30′39″~112°33′41″ E,23°09′21″~23°11′30″ N),受南亞熱帶季風(fēng)濕潤(rùn)氣候影響,年平均氣溫為20.9℃,其中,最冷月1月平均氣溫為12.0℃,最熱月7月平均氣溫為28.0℃,全年總降水量達(dá)1 956 mm并有明顯的干濕季,濕季主要集中在4-9月, 干季主要集中在10月到次年3月,年均相對(duì)濕度為80.8%,年均蒸發(fā)量為1 115 mm[16]。鼎湖山主要植被類型為季風(fēng)常綠闊葉樹(shù)、針闊葉混交林和馬尾松林。本研究樣地設(shè)置在針闊葉混交林中, 研究對(duì)象為錐栗,土壤類型為強(qiáng)酸性土,土壤有機(jī)質(zhì)含量與土壤養(yǎng)分含量都較高[17]。
2 材料和方法
2.1 野外采樣和氣象數(shù)據(jù)獲取
在鼎湖山針闊葉混交林選擇3個(gè)采樣點(diǎn)(圖1),海拔分別為58 (樣點(diǎn)1)、214 (樣點(diǎn)2)和333 m (樣點(diǎn)3)。每個(gè)采樣點(diǎn)選擇3~5棵生長(zhǎng)狀況良好且無(wú)病蟲(chóng)害的錐栗為研究對(duì)象,從2015年年初開(kāi)始,每2周用微采樣技術(shù)進(jìn)行1次樹(shù)木微樣芯采集,方法是利用微型生長(zhǎng)錐(trephor)在樹(shù)干離地1~1.3 m的高度沿螺旋上升進(jìn)行采集[18],2015年野外總共采集到325個(gè)錐栗微樹(shù)芯樣本。同時(shí), 通過(guò)鼎湖山的氣象觀測(cè)站(112°30′39″~112°33′41″E,23°09′21″~23°11′30″ N)獲取2015年樣地的日最高溫(℃)、日最低溫(℃)、日均相對(duì)濕度(%)、日降雨量(mm)以及光合有效輻射[μmol/(m2·s)]等氣象數(shù)據(jù)。
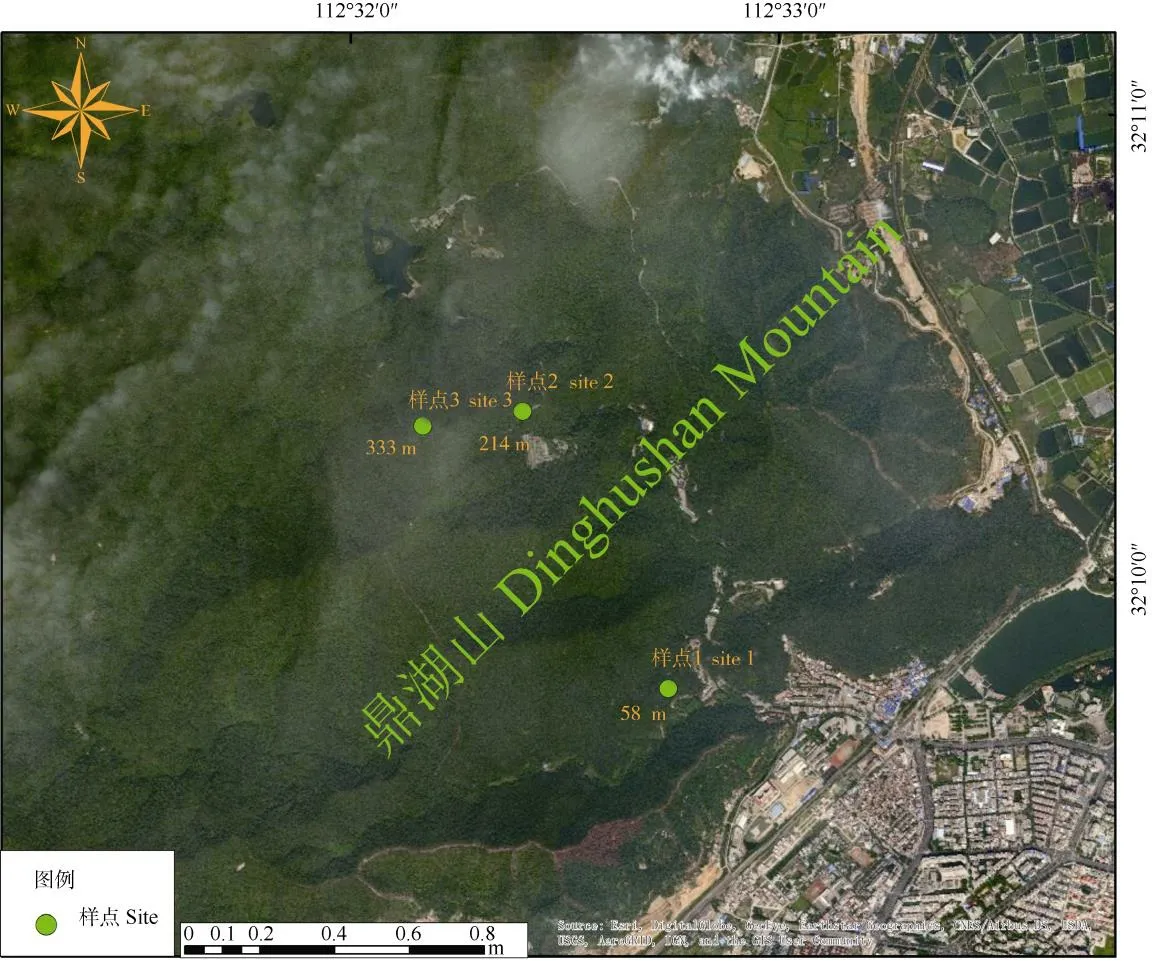
圖1 試驗(yàn)樣地地理位置示意圖Fig. 1 Location of experimental sites
2.2 微樹(shù)芯樣本處理
將野外采集的樣本先放入軟化劑(體積比為甘油∶無(wú)水乙醇∶蒸餾水=1∶2∶3)中軟化,然后逐一放入90%乙醇(1.5 h)、90%乙醇(1.5 h)、95%乙醇(1 h)、無(wú)水乙醇(1 h)、無(wú)水乙醇(1 h)中脫水;再放入純檸檬烯溶液中透明處理3次,每次處理2 h。將透明樣品放入純石蠟溶液2 h,共3次,使石蠟完全取代樣本中的透明劑,最后經(jīng)過(guò)包埋、冷凍,在輪轉(zhuǎn)切片機(jī)上切割并制成8~10μm厚的切片。將干燥后的切片依次放入純檸檬烯(共2次)和無(wú)水乙醇(共2次)中脫蠟,每次20 min;然后切片用焦油紫染液進(jìn)行染色(體積比為0.6%焦油紫染液∶蒸餾水=1∶1),置于顯微鏡下觀察,利用LAS V 4.6軟件測(cè)量木質(zhì)部的徑向生長(zhǎng)長(zhǎng)度,觀測(cè)的指標(biāo)為木質(zhì)部年內(nèi)形成總長(zhǎng)度,包含增大期、增厚期和成熟期3個(gè)時(shí)期細(xì)胞的總徑向生長(zhǎng)長(zhǎng)度。每個(gè)樣片沿木質(zhì)部形成的徑向方向隨機(jī)重復(fù)測(cè)量3次并取平均值,減小因測(cè)量產(chǎn)生的誤差[19]。
2.3 數(shù)據(jù)處理
傳統(tǒng)的Gompertz函數(shù)能較好地?cái)M合木質(zhì)部的“S”型生長(zhǎng)動(dòng)態(tài),對(duì)非“S”型的木質(zhì)部生長(zhǎng)動(dòng)態(tài)擬合程度較低,而廣義可加模型(GAMs)是數(shù)據(jù)驅(qū)動(dòng)的模型,可以更好地對(duì)原始數(shù)據(jù)進(jìn)行擬合[9]。因此本研究選用GAMs來(lái)擬合木質(zhì)部的形成動(dòng)態(tài)。GAMs方程為:y=f(w), 式中,y為響應(yīng)變量(新形成的木質(zhì)部每?jī)芍艿纳L(zhǎng)長(zhǎng)度),w為木質(zhì)部寬度即生長(zhǎng)量,f為平滑函數(shù)。
為進(jìn)一步了解錐栗年內(nèi)生長(zhǎng)狀況,計(jì)算錐栗的生長(zhǎng)速率,即每棵樹(shù)每2周木質(zhì)部的生長(zhǎng)速率:Ri=Wi-Wi-2, 式中,R為第i周木質(zhì)部生長(zhǎng)速率;Wi和Wi-2分別為當(dāng)周和其前兩周木質(zhì)部的生長(zhǎng)量。本研究根據(jù)生長(zhǎng)速率的峰值,將生長(zhǎng)季分為生長(zhǎng)季前期和生長(zhǎng)季后期兩個(gè)時(shí)間段。生長(zhǎng)季前期為生長(zhǎng)開(kāi)始到最大生長(zhǎng)率的日期,生長(zhǎng)季后期定義為最大生長(zhǎng)速率日期到生長(zhǎng)季結(jié)束。
首先,用混合模型分析樹(shù)木生長(zhǎng)動(dòng)態(tài)是否在不同樣點(diǎn)間存在顯著差異,Yije=α+β1Sij+εij, 其中,Yij是樣點(diǎn)e第i棵樹(shù)在第j周的木質(zhì)部生長(zhǎng)速率,α是截距,S為固定因子樣點(diǎn),εij是誤差項(xiàng)。然后,計(jì)算采樣前兩周對(duì)應(yīng)的平均溫度、累計(jì)降雨量、相對(duì)濕度、累計(jì)有效輻射,并采用混合模型探索木質(zhì)部生長(zhǎng)速率與對(duì)應(yīng)時(shí)間段氣候因子的關(guān)系。考慮到環(huán)境因子間的共線性,本研究建立了4個(gè)模型,分別探索全年尺度上平均溫度、累計(jì)降雨量、相對(duì)濕度和累計(jì)光合有效輻射對(duì)木質(zhì)部生長(zhǎng)的影響,Yij=α+β1Xij+εij, 其中,Yij是第i棵樹(shù)在第j周的木質(zhì)部生長(zhǎng)速率,α是截距,Xij代表第i棵樹(shù)在第j周的氣候因子,εij是誤差項(xiàng)。同時(shí),用同樣的方法探索在生長(zhǎng)季前期和后期,木質(zhì)部生長(zhǎng)與氣候之間的關(guān)系。
用R 3.5.0進(jìn)行數(shù)據(jù)處理并采用sigmaplot 13.0軟件畫(huà)圖。
3 結(jié)果和分析
3.1 木質(zhì)部年內(nèi)生長(zhǎng)動(dòng)態(tài)
通過(guò)鏡下觀察切片,錐栗木質(zhì)部生長(zhǎng)的起始時(shí)間早于觀測(cè)的時(shí)間,即在2015年doy70 (70 days of year) (3月初)木質(zhì)部已經(jīng)開(kāi)始生長(zhǎng)。3個(gè)樣點(diǎn)監(jiān)測(cè)的目標(biāo)樹(shù)木質(zhì)部的年內(nèi)生長(zhǎng)動(dòng)態(tài)均顯示,錐栗木質(zhì)部均持續(xù)緩慢生長(zhǎng)(圖2),其中樣點(diǎn)1的木質(zhì)部生長(zhǎng)開(kāi)始時(shí)間最早,約為2015年的doy70 (3月初), 樣點(diǎn)2的稍晚于樣點(diǎn)1,約在2015年doy90 (3月底),樣點(diǎn)3的最晚,約在doy100 (4月初)。
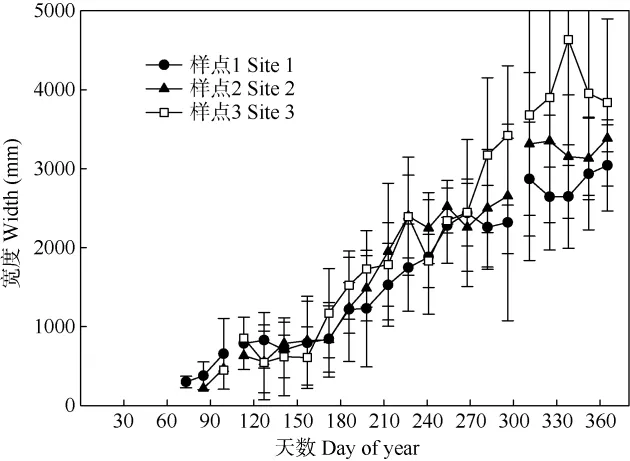
圖2 2015年3個(gè)樣點(diǎn)錐栗木質(zhì)部的累積生長(zhǎng)動(dòng)態(tài)Fig. 2 Dynamic changes in annual cumulative growth of Castanea henryi xylem at three sites in 2015
混合模型分析表明,樣點(diǎn)作為固定因子不顯著,說(shuō)明樹(shù)木生長(zhǎng)狀況在不同的海拔沒(méi)有顯著差異。通過(guò)GAMs擬合樹(shù)木的木質(zhì)部生長(zhǎng)動(dòng)態(tài),擬合精度高達(dá)95%。鼎湖山錐栗木質(zhì)部在doy70左右開(kāi)始迅速生長(zhǎng),在doy100~doy150的生長(zhǎng)速率趨于平緩,doy150后錐栗再次迅速生長(zhǎng),直至2015年12月底依舊緩慢生長(zhǎng)(圖3)。
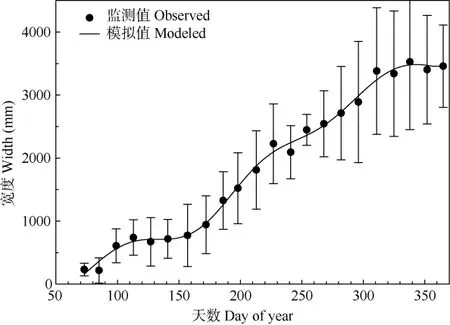
圖3 GAMs擬合的錐栗木質(zhì)部2015年內(nèi)的累積生長(zhǎng)Fig. 3 Annual cumulative growth of Castanea henryi xylem in 2015 with GAMs fit
用GAMs模型擬合錐栗生長(zhǎng)速率,擬合度較低,僅為50%,這可能是由于樹(shù)木之間的生長(zhǎng)速率差異較大導(dǎo)致的。盡管如此,仍然可以看到錐栗木質(zhì)部生長(zhǎng)速率呈現(xiàn)明顯的季節(jié)性差異。在doy70~doy200期間,木質(zhì)部迅速生長(zhǎng),在200 d左右生長(zhǎng)速度達(dá)到最大值,而后逐漸趨于平緩,300 d之后下降,但仍以較緩慢的速度生長(zhǎng)(圖4)。因此本研究中將doy200作為一個(gè)分界點(diǎn),將doy200之前界定為錐栗木質(zhì)部生長(zhǎng)季前期,doy200之后界定為錐栗木質(zhì)部生長(zhǎng)季后期。
3.2 氣候因子
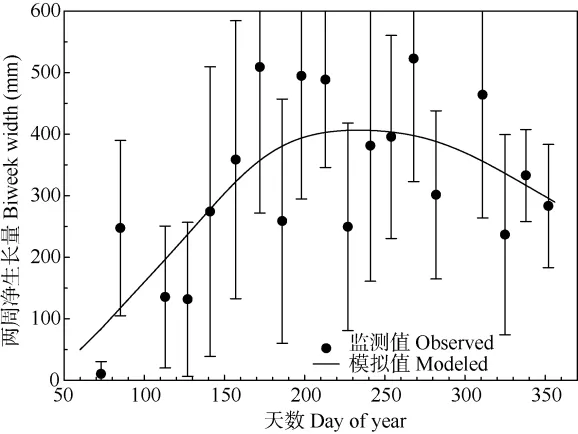
圖4 GAMs擬合的錐栗木質(zhì)部2015年內(nèi)生長(zhǎng)速率Fig. 4 Annual growth rate of Castanea henryi xylem in 2015 with GAMs fit
2015年的樣地氣象數(shù)據(jù)表明(圖5),溫度和光合有效輻射呈現(xiàn)明顯的先升后降的變化趨勢(shì)。doy150之后的溫度、降水都增加。2015年夏季最高溫在7月1日(doy182),達(dá)38.3℃; 全年降雨量有明顯的干濕季之分,濕季集中在4-9月, 10月4日的降雨量最大,為120.4 mm;年平均相對(duì)濕度為78.8%;6月20日的光合有效輻射最高, 為50.902μmol/(m2·s)。
3.3 木質(zhì)部徑向生長(zhǎng)對(duì)氣候因子的響應(yīng)
通過(guò)混合模型分析錐栗木質(zhì)部生長(zhǎng)速率在整年尺度和生長(zhǎng)季早期和晚期對(duì)氣候的響應(yīng)關(guān)系。結(jié)果表明(表1),在整年尺度上,木質(zhì)部生長(zhǎng)速率與日照時(shí)長(zhǎng)存在極顯著的相關(guān)關(guān)系(P<0.001),與平均溫度存在顯著的相關(guān)關(guān)系。從表2可見(jiàn),生長(zhǎng)季前期錐栗木質(zhì)部生長(zhǎng)速率與平均溫度、光合有效輻射均存在極顯著的相關(guān)關(guān)系(P<0.001);而生長(zhǎng)季后期的木質(zhì)部生長(zhǎng)速率與平均溫度、總降水量、年均相對(duì)濕度和有效光合輻射均不存在顯著的相關(guān)關(guān)系(表3)。
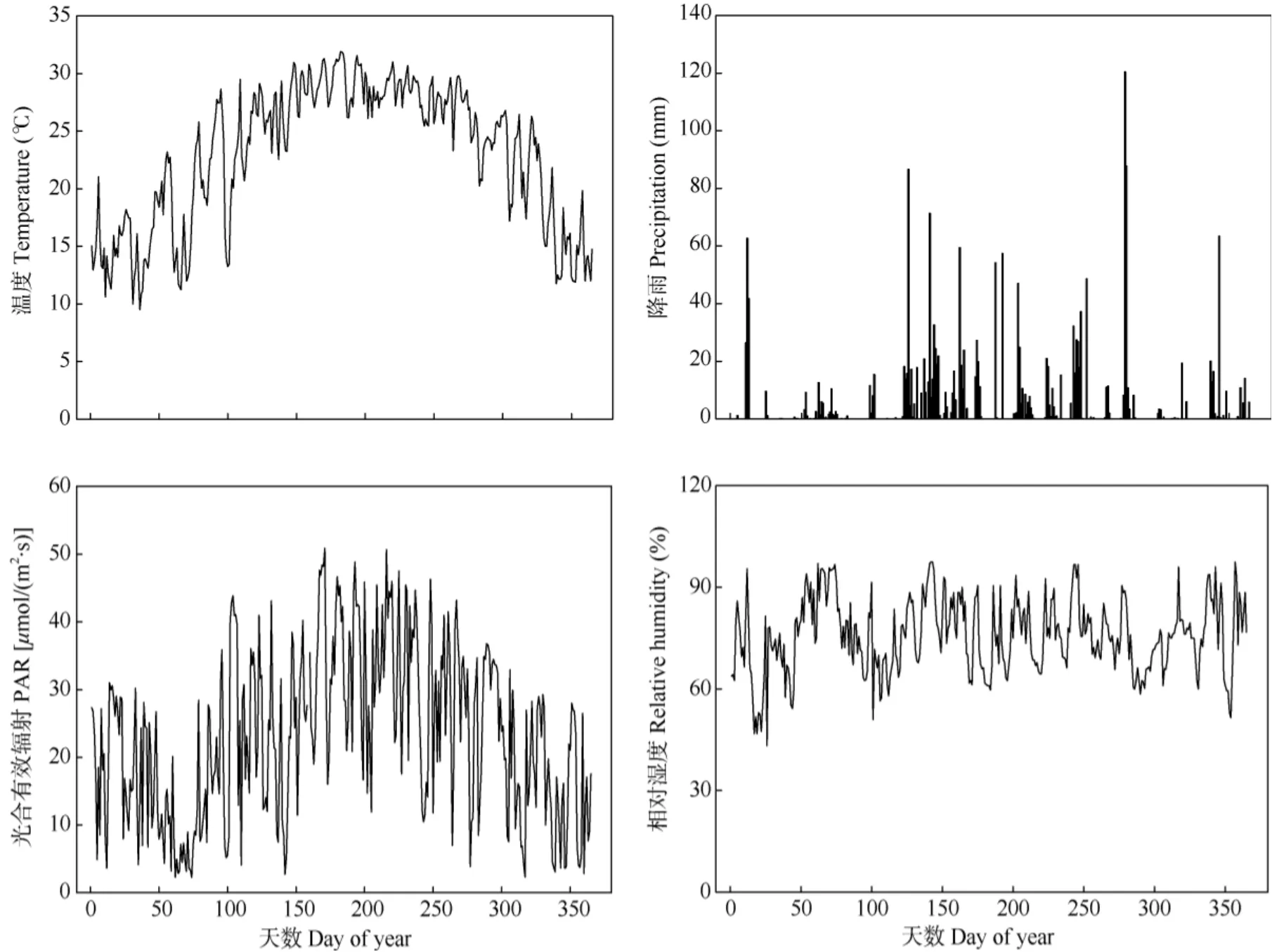
圖5 2015年鼎湖山樣地氣候信息Fig. 5 Climate information of Dinghu Mountain in 2015
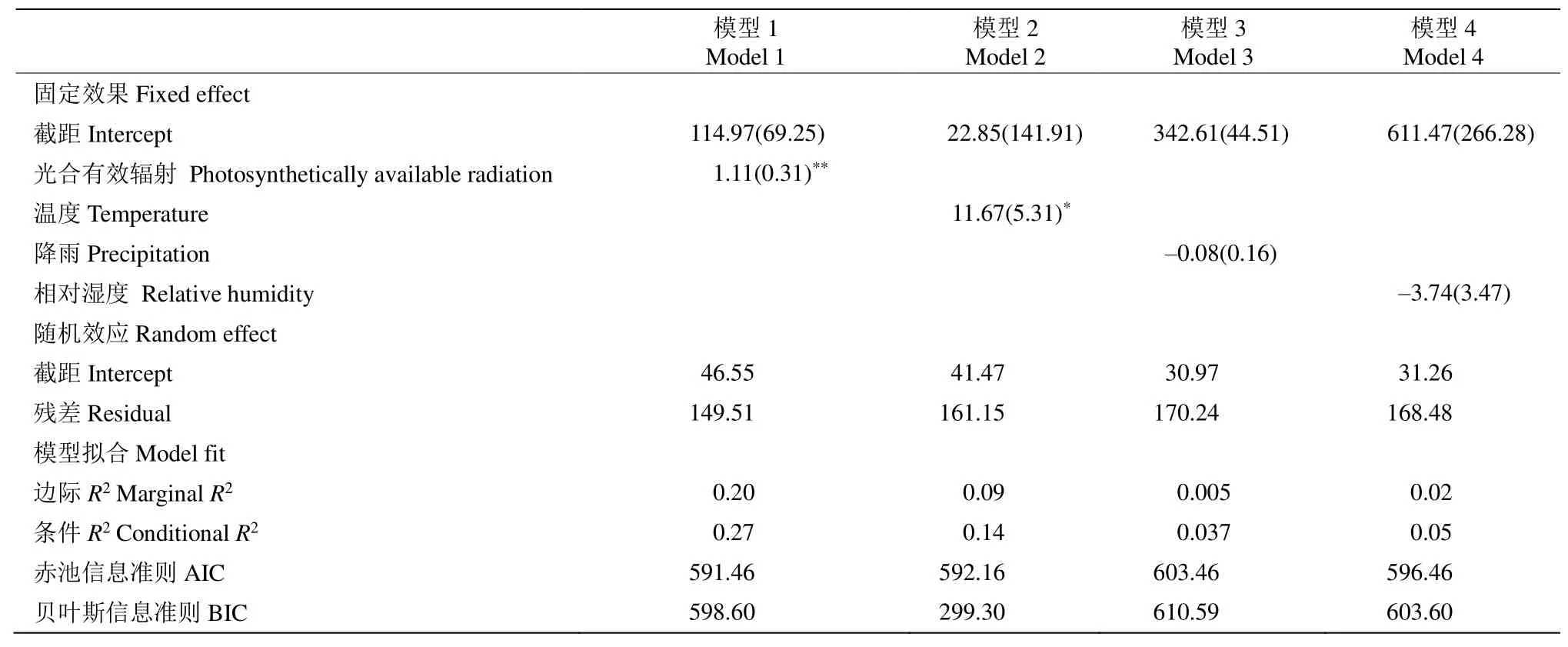
表1 利用混合模型分析錐栗木質(zhì)部年內(nèi)生長(zhǎng)速率與氣候因子的關(guān)系Table 1 Relationship between annual xylem growth rate of Castanea henryi and climate factors by using mixed-effect models
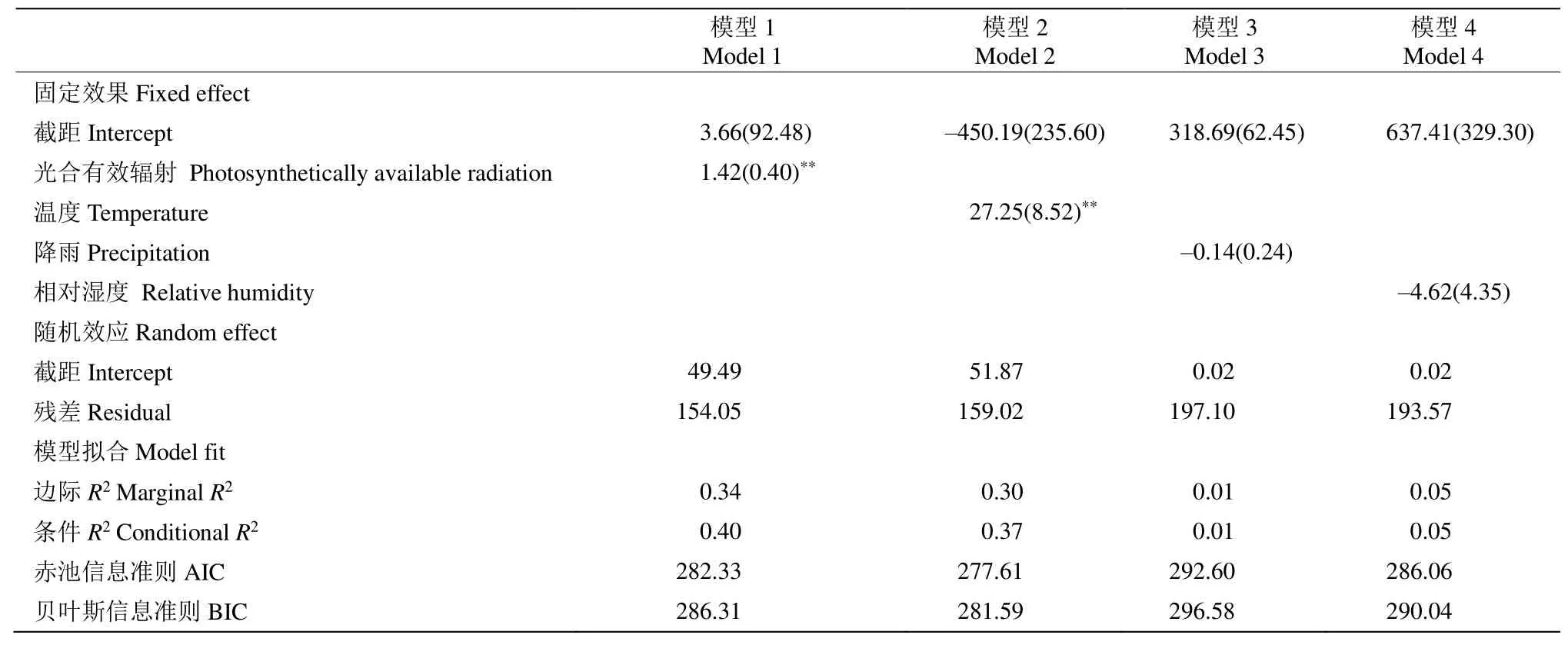
表2 利用混合模型分析錐栗木質(zhì)部生長(zhǎng)季前期生長(zhǎng)速率與氣候因子的關(guān)系Table 2 Relationship between xylem growth rate of Castanea henryi in early growth season and climate factors by using mixed-effect models
4 結(jié)論和討論
4.1 木質(zhì)部年內(nèi)生長(zhǎng)動(dòng)態(tài)
樹(shù)木木質(zhì)部的生長(zhǎng)量由生長(zhǎng)季長(zhǎng)度和生長(zhǎng)速率共同決定, 即使是有著相同的生長(zhǎng)量,也可能有不同的生長(zhǎng)機(jī)制,較短的生長(zhǎng)季和較快的生長(zhǎng)速率或較長(zhǎng)的生長(zhǎng)季和較慢的生長(zhǎng)速率可以產(chǎn)生相同的生長(zhǎng)量。大量研究表明,在溫帶和北寒帶地區(qū), 樹(shù)木木質(zhì)部生長(zhǎng)季一般為4-9月,木質(zhì)部年內(nèi)形成動(dòng)態(tài)呈“S”型生長(zhǎng)曲線[7,20]。相比于溫帶和寒帶,我國(guó)亞熱帶全年溫度較高且降雨充沛,因此樹(shù)木生長(zhǎng)季長(zhǎng)度更長(zhǎng)。本研究中,錐栗的徑向生長(zhǎng)開(kāi)始時(shí)間早于3月,12月底仍保持一定速率的生長(zhǎng),呈現(xiàn)出一種幾乎全年生長(zhǎng)的模式。這和當(dāng)?shù)氐尼樔~樹(shù)馬尾松(Pinus massoniana)相似,驗(yàn)證了中國(guó)亞熱帶地區(qū)樹(shù)木均具有較長(zhǎng)的生長(zhǎng)季的結(jié)論。生長(zhǎng)季的延長(zhǎng)對(duì)木質(zhì)部的生長(zhǎng)量有促進(jìn)作用[21],從而可以進(jìn)一步促進(jìn)森林的固碳作用。
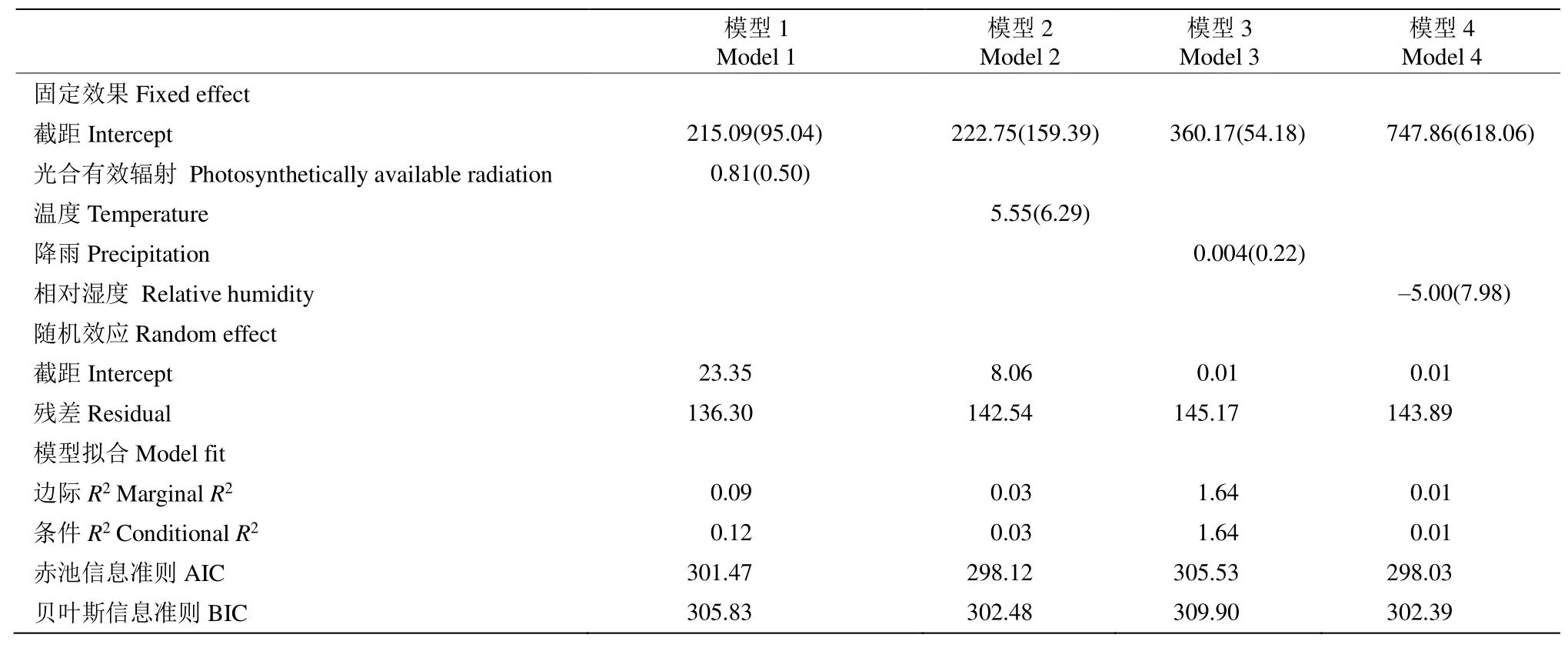
表3 利用混合模型分析錐栗木質(zhì)部生長(zhǎng)季后期生長(zhǎng)速率與氣候因子的關(guān)系Table 3 Relationship between xylem growth rate of Castanea henryi in late growth season and climate factors by using mixed-effect models
之前的研究對(duì)生長(zhǎng)季長(zhǎng)度關(guān)注較多,然而最新研究表明,生長(zhǎng)速率是最后生長(zhǎng)的最終決定因素。通過(guò)敏感性分析探索青藏高原海拔梯度上祁連圓柏(Juniperus przewalskii)木質(zhì)部的生長(zhǎng)物候期,表明木質(zhì)部生長(zhǎng)和生長(zhǎng)速率以及生長(zhǎng)季長(zhǎng)度相關(guān),而生長(zhǎng)速率起到?jīng)Q定作用[22]。木質(zhì)部的生長(zhǎng)速率具有較強(qiáng)的彈性,可以通過(guò)調(diào)節(jié)使樹(shù)木適應(yīng)環(huán)境的變化。本研究中,木質(zhì)部的生長(zhǎng)速率呈現(xiàn)顯著的季節(jié)性差異,并且對(duì)氣候因子的響應(yīng)也不同,說(shuō)明亞熱帶樹(shù)木木質(zhì)部的生長(zhǎng)動(dòng)態(tài)具有較強(qiáng)的靈活性。
之前的研究表明,不同海拔梯度引發(fā)的溫度差異,會(huì)引發(fā)樹(shù)木木質(zhì)部生長(zhǎng)差異[22]。然而,本研究中3個(gè)海拔的錐栗生長(zhǎng)季長(zhǎng)度和生長(zhǎng)量均無(wú)顯著差異,這可能是由于本研究的海拔差異較小,約為100 m,并不能引起樹(shù)木木質(zhì)部生長(zhǎng)模式和生長(zhǎng)量的改變。
4.2 木質(zhì)部生長(zhǎng)對(duì)氣候的季節(jié)性響應(yīng)差異
木質(zhì)部的形成是一個(gè)很復(fù)雜的過(guò)程,既受內(nèi)因的控制(遺傳基因、個(gè)體的發(fā)育狀況、碳水化合物、激素等)[23],也受到外部環(huán)境的影響,如溫度,降雨,光合有效輻射等[5]。同時(shí),這些因子對(duì)樹(shù)木的生長(zhǎng)具有滯后效應(yīng),木質(zhì)部生長(zhǎng)及其形成過(guò)程都取決于當(dāng)年及生長(zhǎng)季前期的許多氣候因子(如溫度、降水、光照時(shí)長(zhǎng)和CO2濃度等)的影響,且這種影響對(duì)木質(zhì)部的形成及樹(shù)木生長(zhǎng)是十分重要的[24]。這種滯后效應(yīng)大多數(shù)是通過(guò)影響可溶性碳水化合物的積累,從而影響樹(shù)木生長(zhǎng),說(shuō)明碳水化合物對(duì)樹(shù)木生長(zhǎng)具有重要的調(diào)節(jié)作用[25]。由于氣候因子和樹(shù)木內(nèi)在因子處于波動(dòng)狀態(tài),樹(shù)木的生長(zhǎng)動(dòng)態(tài)也表現(xiàn)出不同的季節(jié)性差異。大量的研究表明,生長(zhǎng)季開(kāi)始時(shí)期主要受到外在因子的影響,而生長(zhǎng)季結(jié)束時(shí)期更多受到內(nèi)在因子的影響。Luo等[26]通過(guò)同步監(jiān)測(cè)急尖長(zhǎng)苞冷杉(Abies georgeivar.smithii)和方枝柏(Juniperus saltuaria)的徑向生長(zhǎng)和植物氮循環(huán)動(dòng)態(tài),認(rèn)為在夏至日左右,植物通過(guò)調(diào)節(jié)葉片營(yíng)養(yǎng)的周轉(zhuǎn),從而使得氮利用效率和光合效率達(dá)到最大值;同時(shí), 在生長(zhǎng)季前期,樹(shù)木的徑向生長(zhǎng)受到溫度的限制作用;而在生長(zhǎng)季后期,這種限制作用減弱甚至消失。這表明樹(shù)木可以通過(guò)生物鐘的作用,使植物內(nèi)部的資源循環(huán)和外界環(huán)境相匹配,而達(dá)到最優(yōu)化生長(zhǎng)。地中海地區(qū)夏季干旱時(shí),植物通過(guò)落葉來(lái)降低水分消耗,并且形成層停止分裂;當(dāng)水分條件滿足時(shí), 形成層開(kāi)始二次分裂,這種“雙峰型”的木質(zhì)部形成動(dòng)態(tài)也展現(xiàn)了明顯的季節(jié)差異,顯示了樹(shù)木生長(zhǎng)對(duì)環(huán)境的適應(yīng)[27]。因此,我們應(yīng)更加深入研究樹(shù)木生長(zhǎng)的季節(jié)性差異,從而全面了解樹(shù)木的生長(zhǎng)機(jī)制以及對(duì)環(huán)境的適應(yīng)。
4.3 生長(zhǎng)季前期生長(zhǎng)機(jī)制
在溫帶和北方森林,普遍認(rèn)為溫度是控制春季樹(shù)木徑向生長(zhǎng)的重要因素[19],然而溫度對(duì)樹(shù)木生長(zhǎng)的促進(jìn)作用是非線性的,當(dāng)溫度超過(guò)一定閾值,樹(shù)木的呼吸作用也會(huì)增強(qiáng),從而導(dǎo)致累積的光合產(chǎn)物下降,進(jìn)而抑制樹(shù)木生長(zhǎng)。在樹(shù)木生長(zhǎng)季早期,溫度對(duì)樹(shù)木徑向開(kāi)始生長(zhǎng)有著積極的促進(jìn)作用[28]。興安落葉松(Larix gmelinii)與氣候因子的響應(yīng)研究表明,5月氣溫升高有利于樹(shù)木的發(fā)芽,使落葉松葉片快速生長(zhǎng),進(jìn)而促進(jìn)木質(zhì)部的生長(zhǎng),易形成寬輪[29]。本研究結(jié)果驗(yàn)證了溫度對(duì)樹(shù)木生長(zhǎng)的促進(jìn)作用,說(shuō)明即使在亞熱帶,全年溫度較高,但在生長(zhǎng)季早期,溫度仍然是限制因子,適宜溫度的提高能顯著促進(jìn)樹(shù)木生長(zhǎng)。
較長(zhǎng)的日照時(shí)間意味著較長(zhǎng)的光合作用時(shí)間,能顯著促進(jìn)光合產(chǎn)物的積累,從而促進(jìn)樹(shù)木生長(zhǎng)。大量研究表明,光照對(duì)木質(zhì)部細(xì)胞的生成量有著重要的影響。歐洲山毛櫸(Fagus sylvatica)的光合速率在光照時(shí)長(zhǎng)最長(zhǎng)的6月達(dá)到最大值,從而顯著促進(jìn)了木質(zhì)部細(xì)胞的生長(zhǎng)[30]。Huang等[31]對(duì)中國(guó)南亞熱帶馬尾松木質(zhì)部的形成進(jìn)行了連續(xù)監(jiān)測(cè),樹(shù)木在干季(9-11月)的生長(zhǎng)明顯比濕季更旺盛,表明在水熱充足的亞熱帶,馬尾松的生長(zhǎng)更多是受凈光合能量的影響。本研究結(jié)果與其一致,說(shuō)明在亞熱帶,溫度和光周期決定的光合產(chǎn)物積累為木質(zhì)部生長(zhǎng)提供了重要的能量,進(jìn)一步?jīng)Q定了木質(zhì)部生長(zhǎng)。
水分可以通過(guò)維持細(xì)胞膨壓來(lái)參與植物的生理活動(dòng),從而調(diào)節(jié)樹(shù)木的生長(zhǎng)。Deslauriers等[32]報(bào)道水分是調(diào)節(jié)黑云杉(Picea mariana)幼苗樹(shù)干形成層活動(dòng)的主要限制因子。然而,本研究結(jié)果表明錐栗年內(nèi)木材的生長(zhǎng)速率與降雨量或相對(duì)濕度之間沒(méi)有顯著的相關(guān)關(guān)系,這表明降水不是鼎湖山錐栗生長(zhǎng)的限制因素,這可能是由于亞熱帶的濕季降雨充足,充裕的降水滲透到土壤底層,即使到了干季,也可以被樹(shù)木根系利用;另外,亞熱帶的樹(shù)木已經(jīng)充分適應(yīng)了當(dāng)?shù)丨h(huán)境,在干季可以通過(guò)調(diào)節(jié)蒸騰作用等來(lái)控制水分的輸出,從而保證樹(shù)木生長(zhǎng)所需水分。
4.4 生長(zhǎng)季后期生長(zhǎng)機(jī)制
在生長(zhǎng)季后期,氣候因子對(duì)樹(shù)木生長(zhǎng)沒(méi)有顯著影響,說(shuō)明木質(zhì)部生長(zhǎng)很可能受到內(nèi)在因子(如光合作用產(chǎn)物、碳水化合物等)的調(diào)控。一般而言,樹(shù)木的碳匯呈現(xiàn)出季節(jié)性動(dòng)態(tài),在生長(zhǎng)季早期,樹(shù)木通過(guò)光合作用固定的碳用于樹(shù)木生長(zhǎng);而在生長(zhǎng)季后期,光合作用固定的碳開(kāi)始積累,用于樹(shù)木下一年的生長(zhǎng),尤其對(duì)于環(huán)孔材,次年春季需要產(chǎn)生大導(dǎo)管,從而完成水分運(yùn)輸?shù)墓δ堋6髮?dǎo)管的產(chǎn)生需要消耗大量能量,春季剛開(kāi)始,葉片光和能力較弱,因此,樹(shù)木的生長(zhǎng)主要依賴于上一年積累的碳水化合物[9,33]。因此對(duì)于樹(shù)木而言,即使有充足的碳匯,碳的可利用性也受到樹(shù)木的發(fā)育階段和環(huán)境因子的限制[34-35]。Huang等[9]對(duì)加拿大魁北克北部針葉林對(duì)香脂冷杉(Abies balsamea)和黑云杉進(jìn)行連續(xù)監(jiān)測(cè)(2010-2013年),認(rèn)為針葉發(fā)芽之前木質(zhì)部形成的主要?jiǎng)恿?lái)源是上一年儲(chǔ)存的能量和碳,而針葉發(fā)芽后新合成的光合作用產(chǎn)物才是木質(zhì)部生長(zhǎng)所需的能量來(lái)源。本研究中,生長(zhǎng)季后期木質(zhì)部生長(zhǎng)速率與氣候因子之間的相關(guān)關(guān)系并不顯著,說(shuō)明生長(zhǎng)季后期光合作用合成的碳一部分用于樹(shù)木木質(zhì)部的生長(zhǎng),而另一部分用于儲(chǔ)存,以供次年樹(shù)木的生長(zhǎng)。因此,這一機(jī)制導(dǎo)致與光合作用直接相關(guān)的氣候因子對(duì)樹(shù)木生長(zhǎng)的影響不顯著。碳水化合物作為樹(shù)木生長(zhǎng)的重要因子,其季節(jié)性動(dòng)態(tài)對(duì)于樹(shù)木發(fā)育有調(diào)節(jié)作用,應(yīng)加強(qiáng)該方面的研究,從而更好地揭示樹(shù)木生長(zhǎng)的內(nèi)在調(diào)節(jié)機(jī)制。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
