木芙蓉品種“單瓣紅”種子萌發特性研究
2020-10-09 11:30:34楊苑釗曾心美馬嬌石小慶朱章順李方文
湖南農業科學 2020年8期
楊苑釗 曾心美 馬嬌 石小慶 朱章順 李方文

摘 要:為探究不同處理對木芙蓉品種“單瓣紅”種子萌發的影響及其對干旱、鹽堿脅迫的響應,采用機械損傷、濃硫酸浸種、赤霉素(GA3)浸種及復合處理破除種子休眠,并采用不同濃度PEG6000和NaCl處理種子后測定種子發芽指標。結果表明:機械損傷15 min和濃硫酸浸種15 min處理組合對于打破種子休眠的效果最佳,種子萌發率達到90%以上;種子萌發與PEG6000和NaCl的脅迫濃度呈線性相關關系,種子發芽率、發芽勢均隨PEG6000和NaCl濃度增加而降低,在低濃度PEG6000和NaCl處理條件下,種子萌發指標與對照差異不顯著,“單瓣紅”種子能夠在中度鹽堿脅迫下萌發、生長,其對NaCl的中耐受濃度為6.7 g/L。
關鍵詞:木芙蓉;單瓣紅;種子;萌發;脅迫
中圖分類號:S685.99文獻標識碼:A文章編號:1006-060X(2020)08-0016-05
Abstract: In this paper, abrading seed coat, seed-soaking with sulphuric acid and gibberellin solutions, and the corresponding combined treatments were used to break seed dormancy to explore their influences on seed germination of the Hibiscus mutabilis L. cultivar “Danbanhong”. In addition, the responses of “Danbanhong” seeds to simulated drought and saline-alkali stresses were studied by soaking seeds with different levels of PEG6000 and Nacl solutions, and then seed germination indices were measured. ?The results showed that ?abrading seed coat for 15 min combined with ?soaking seeds in sulphuric acid solution for 15min was the best for seed dormancy break, and the germination percentage was more than 90%. There was a linear correlation between the germination percentage and the concentrations of PEG6000 and NaCl solutions. The germination percentage and germination energy decreased with the increase of PEG 6000 and NaCl concentrations. The seed germination indices under low concentrations of PEG6000 and NaCl had no significant difference in comparison with those of CK (15-min abrading seed coat + 15-min soaking seeds in sulphuric acid solution). “Danbanhong” seeds can germinate and grow under moderate saline-alkali stress with a Macl-tolerant concentration of 6.7 g/L.
Key words: Hibiscus mutabilis L.; Danbanhong; seed; germination; stress
木芙蓉(Hibiscus mutabilis L.)別名拒霜花,為錦葵科木槿屬花卉,原產于中國,在東亞及東南亞地區均有分布。木芙蓉花大色艷,花期較長,在成都地區栽植較為廣泛[1]。種子休眠是指具有正常活力的種子在適宜的環境條件下仍然不能萌發的現象[2]。植物種子的休眠與其遺傳特性相關,但外界環境因素在很大程度上也會影響種子的休眠特性[3]。植物種子的休眠能夠幫助植物度過逆境,因此休眠對于植物適應環境具有重要的生態意義[4]。但與此同時,植物的休眠也給品種培育及應用帶來諸多不便和困難,嚴重影響園林景觀植物的豐富性。因此,研究如何打破園林景觀植物種子的休眠非常重要。如何打破種子休眠一直是種子科學的研究熱點[5]。破除種子休眠的方法主要有物理、化學、生物和綜合法[6-7]。土壤干旱和鹽堿化是制約種子萌發的主要影響因素[8]。He等[9]研究預測未來環境惡化程度將會加劇。木芙蓉作為成都市市花,觀賞價值極高,但對于如何打破木芙蓉種子休眠及其與干旱脅迫和鹽堿脅迫的相關性研究鮮有報道。為了進一步優化打破木芙蓉種子休眠的方法,并探討木芙蓉種子對于干旱和鹽堿的適應性,筆者以木芙蓉品種“單瓣紅”種子為材料,研究了采用濃硫酸浸種、機械損傷、GA3浸種等方式打破種子休眠的效果,同時使用聚乙二醇(PEG6000)和NaCl溶液模擬干旱和鹽脅迫,對木芙蓉種子萌發的抗旱抗鹽特征進行研究。
1 材料與方法
1.1 試驗材料
木芙蓉“單瓣紅”種子(千粒重4.70±0.21 g)于2018年11月在成都市植物園苗圃采集。種子存放于干燥通風處,于2019年3—10月在成都市植物園開展試驗。試驗所用PEG6000和NaCl均為分析純。
1.2 試驗設計
1.2.1 種子預處理 選取飽滿健壯,大小均一,無病蟲為害的種子,用1%高錳酸鉀消毒20 min,用清水洗凈后待用。
1.2.2 打破種子休眠方法 打破種子休眠方法試驗包括:機械破損、硫酸浸種、GA3/丙酮浸種及不同因素的復合處理,共4個試驗。
(1)機械損傷試驗。用砂紙磨擦種子,擦破種皮,但不傷及其胚,磨擦時間設5、10、15、20 min 4個處理,每個處理30粒種子,重復3次。
(2)硫酸浸種試驗。用98%的濃硫酸浸種,并不停攪拌。浸種時間設5、10、15、20 min 4個處理,處理完后,用清水將種子洗凈,pH試紙檢測呈中性后晾干備用,每個處理30粒種子,重復3次。
(3)GA3/丙酮浸種試驗。以丙酮為溶劑,配制100 mg/L的GA3溶液,用該溶液浸種。浸種時間設3、6、12、24 h 4個處理,處理完后將種子洗凈備用,每個處理30粒種子,重復3次。
(4)復合處理試驗。包括下列3個處理。機械損傷+硫酸浸種:選用(1)和(2)中發芽率最高的處理水平進行復合處理。機械損傷+GA3/丙酮浸種:選用(1)和(3)中效果最好的處理水平進行復合處理。硫酸浸種+GA3/丙酮浸種:選用(2)和(3)中效果最好的處理水平進行復合處理。機械損傷+硫酸浸種+GA3/丙酮浸種:選用(1)、(2)和(3)中效果最好的處理水平進行復合處理。每個處理均為30粒種子,重復3次。
上述試驗均以不處理作對照(CK)。將種子置于墊有2層濾紙的培養皿中,放入人工氣候箱中培養。培養條件設置為恒溫20℃,光照12 h,黑暗12 h。每日定期觀察種子萌發情況,“露白”即認定為發芽。
1.2.3 PEG和NaCl脅迫對種子萌發的影響 采用PEG6000和NaCl溶液模擬滲透勢和鹽脅迫。以1.2.2中最佳處理方式作為對照。將“單瓣紅”種子按照1.2.2中最佳處理方式處理后,分別使用不同濃度的PEG6000和NaCl溶液浸種,PEG6000濃度設0、5%、10%、15%、20%、25%、30%共7個處理水平;NaCl濃度設0、0.2%、0.4%、0.6%、0.8%、1.0%共6個處理水平。浸種24 h后置于暗處,瀝干水分后放于培養皿中,置于人工氣候箱中培養。培養條件設置為恒溫20℃,光照12 h,黑暗12 h。每個處理30粒種子,重復3次。每日定期觀察種子萌發情況,“露白”即認定為發芽。脅迫試驗結束后5 d,統計各處理萌發幼苗根長。
1.3 種子萌發指標
統計各處理發芽15 d的發芽情況。分別按公式(1)、(2)、(3)、(4)計算發芽率(GP)、發芽勢(GE)、發芽指數(GI)、平均發芽時間(MGT)。
1.4 數據處理
數據采用Excel 2010軟件進行整理,采用SPSS18.0統計軟件對GP、GE、GI、MGT進行處理和差異顯著性分析(LSD法)。
2 結果與分析
2.1 打破木芙蓉種子休眠的方法研究
2.1.1 機械損傷打破種子休眠的效果 木芙蓉“單瓣紅”種子經機械損傷種皮后其萌發指標均顯著提高(見表1)。種子經機械損傷15 min處理的發芽率、發芽勢和發芽指數分別為40.00%、28.89%和15.36,均顯著高于CK和機械損傷5 min的處理(除機械損傷5 min處理的發芽率外),但與機械損傷10和20 min的處理差異不顯著。平均發芽時間在機械損傷不同時長之間沒有顯著差異。
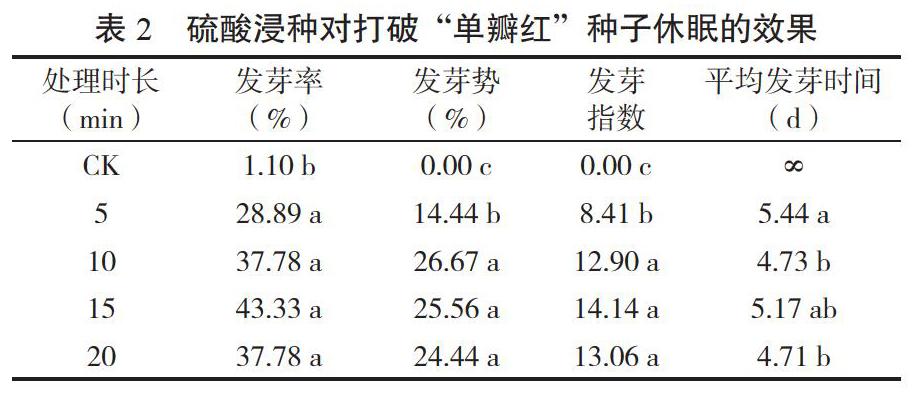
2.1.2 硫酸浸種打破種子休眠的效果 不同硫酸浸種時長對木芙蓉“單瓣紅”種子萌發指標的影響不同(見表2)。未處理(CK)的種子發芽率極低,為1.10%,硫酸浸種處理后的發芽率、發芽勢和發芽指數都顯著高于對照(P<0.05)。其中,硫酸浸種15 min的發芽率最高,為43.33%,為CK發芽率的39.4倍,發芽勢由CK的0顯著提高到了25.56%,發芽指數由CK的0顯著提高到了14.14。浸種5~20 min處理之間的發芽率無顯著性差異,但浸種5 min的發芽勢和發芽指數顯著低于其他時長處理,其他時長處理之間的發芽勢和發芽指數差異不顯著。對照(CK)在試驗結束時種子仍未萌發,故平均發芽時間(MGT)記為∞。
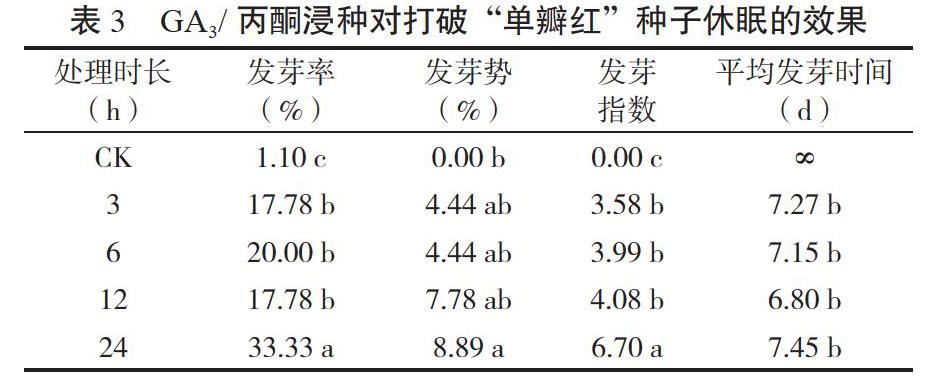
2.1.3 GA3/丙酮浸種打破種子休眠的效果 GA3/丙酮浸種能夠顯著提高發芽率、發芽勢和發芽指數,降低平均發芽時間(見表3)。以GA3/丙酮浸種24 h處理的效果最好,其種子萌發指標均顯著高于對照,且發芽率和發芽指數2個指標均顯著高于其他時長處理,結果與梁靜等[10]的研究結果部分一致,其差異可能由于供試材料和處理時間梯度不同所致。
2.1.4 復合處理打破種子休眠的效果 由圖1可知,機械損傷種皮15 min結合硫酸浸種15 min處理的萌發效果最好,其發芽率、發芽勢和發芽指數分別達到97.78%、97.78%和69.17,顯著高于其他處理和對照,其平均發芽時間為2.00 d,顯著低于其他處理和對照,表明該復合處理為打破種子休眠的最佳條件。
2.2 PEG和NaCl脅迫對木芙蓉種子萌發特性的影響
2.2.1 種子萌發特性對干旱脅迫的響應 由表4可知,“單瓣紅”種子的各項萌發指標都隨著PEG6000的處理濃度提高而降低。5%PEG處理的發芽率、發芽勢和平均發芽時間與對照差異不顯著,PEG濃度達到10%以上時,發芽率、發芽勢和發芽指數都顯著低于對照。當PEG濃度為25%時,發芽率、發芽勢和發芽指數極低,分別僅為1.11%,1.11%和0.70,且有2個重復種子沒有萌發。當PEG濃度為30%時,3個重復種子均沒有萌發。
如圖2所示,隨著PEG6000處理濃度的增加,“單瓣紅”幼苗根長呈現先增加后下降的趨勢。處理濃度為5%時,幼苗根長為71.58 mm,比對照長14.06 mm,說明輕度干旱脅迫能夠促進幼苗根系生長。當處理濃度為10%及以上時,幼苗根長均短于對照,且處理濃度達到25%和30%(此時溶液的滲透勢分別為-0.79和-1.10 Mpa)時,幼苗根長顯著低于其他處理和對照。
2.2.2 種子萌發特性對鹽堿脅迫的響應 根據表5可知,低濃度NaCl脅迫(0.2%~0.4%)對“單瓣紅”種子的萌發指標影響不顯著。在0.6%和0.8%NaCl脅迫下,其發芽率顯著低于對照組,但仍能達到65.56%和55.56%。而在1%NaCl脅迫下的發芽率下降到22.22%,顯著低于CK和其他處理,發芽勢和發芽指數也分別僅為4.44%和3.62,平均發芽時間則高到10.09 d,顯著高于其他處理和對照。
如圖2可示,“單瓣紅”幼苗的根長隨著NaCl溶液濃度的提高而降低。0.2%和0.4%NaCl處理的根長分別為55.61和44.81 mm,與對照差異不顯著。但0.6%及以上濃度處理則會顯著抑制幼苗根的生長。
2.2.3 “單瓣紅”種子對不同脅迫劑的耐受濃度 以脅迫劑的濃度為自變量(x),以種子發芽指數為因變量(y)建立回歸方程(表6)。再以種子發芽指數分別為對照的50%和10%作為“單瓣紅”種子對脅迫劑的半耐受劑量和極限耐受劑量,可以計算出PEG6000的中耐受濃度和極限耐受濃度分別為129和244 g/L;NaCl的中耐受濃度和極限耐受濃度分別為6.7和9.5 g/L。
3 討 論
環境和激素的變化是決定種子休眠與萌發的重要因素。由于種子的種皮致密、堅硬及具有蠟質而導致水分不能進入,也是造成植物種子休眠的主要原因[10]。PEG和NaCl已被廣泛應用于植物的抗旱、抗鹽研究[11]。PEG是一種高分子滲透劑,能夠控制進入種子的水分含量,使植物種子處于類似干旱脅迫之中。低濃度NaCl能夠參與部分酶的激活[12],但高濃度鹽脅迫一方面會對植物造成滲透脅迫引起植物失水,另一方面也會引起植物離子中毒[13]。
試驗采用的打破種子休眠方法均能顯著提高“單瓣紅”種子的發芽率。梁靜等[14]的研究結果表明,機械損傷處理和GA3浸種都能有效打破木芙蓉種子休眠,筆者的研究也得出了同樣的結果。硫酸浸種對打破木芙蓉種子休眠鮮有報道,試驗采用98%濃硫酸浸種后,“單瓣紅”的發芽率呈現先上升后下降的趨勢,浸種15 min為促進種子萌發的最佳浸種時長,但當浸種時間繼續延長時則發芽率開始下降,可能原因是長時間濃硫酸浸泡使種皮崩潰,傷及了種胚[15]。采用機械損傷種皮15 min結合硫酸浸種15 min對打破“單瓣紅”種子休眠的效果最好,該處理的發芽率能夠達到90%以上,可以在生產上應用。
試驗采用聚乙二醇(PEG6000)模擬干旱脅迫,研究其對“單瓣紅”種子萌發影響的結果是:PEG脅迫對種子萌發具有一定的抑制和延緩作用,在低濃度條件下,PEG對種子萌發的延緩和抑制作用不明顯,輕度干旱脅迫(5%PEG)還能促進木芙蓉幼苗根部生長。“單瓣紅”種子萌發這種對干旱脅迫的響應與木荷(Schima superba)、矮沙冬青及馬尾松的響應機制[16-18]相一致。但陳士超等[19]對紫花苜蓿(Medicago sativa)的研究結果表明,隨著PEG濃度的增加,紫花苜蓿種子的萌發指標呈現先上升后下降的趨勢,說明低濃度的PEG脅迫能夠對紫花苜蓿種子產生正向影響,其原因可能是由于不同物種本身的生物學特性對干旱脅迫的響應程度和范圍不同。
試驗表明,低濃度NaCl脅迫對“單瓣紅”種子萌發無顯著抑制作用,這與白菜、紫花苜蓿種子對低鹽堿脅迫的響應[20]一致。當NaCl溶液的濃度提高到0.6%及以上時,對“單瓣紅”種子萌發開始有顯著的抑制作用,但在0.6%的NaCl脅迫下,發芽率仍然達到65.56%,發芽勢和發芽指數分別達到45.56%和32.50,“單瓣紅”種子對NaCl的中耐受濃度為6.7 g/L。
因此,“單瓣紅”種子對中度鹽堿脅迫具有一定的適應性,木芙蓉可以在一定程度上用于鹽堿土壤的改良。但若在鹽堿地區大面積推廣應用木芙蓉,還需對其成株的抗鹽性進行進一步研究和觀察。
參考文獻:
[1] 楊苑釗,曾心美,馬 嬌,等. 木芙蓉不同早花品種花期特征觀察與分析[J]. 現代農業科技,2019(17):144-145,150.
[2] 楊期和,葉萬輝,宋松泉,等. 植物種子休眠的原因及休眠的多形性[J]. 西北植物學報,2003,23(5):837-843.
[3] 傅 強,楊期和,葉萬輝. 種子休眠的解除方法[J]. 基因組學與應用生物學,2003,22(3):230-234.
[4] 陳乙實,娜麗克斯·外里,王樹林,等. 不同處理方法對7種豆科植物種子休眠以及成苗特性的影響[J]. 草地學報,2017,25(4):823-831.
[5] 胡小文,武艷培,王彥榮,等. 豆科種子休眠破除方法初探[J]. 西北植物學報,2009,29(3):568-573.
[6] 李兵兵,魏小紅,徐 嚴. 麻花秦艽種子休眠機理及其破除方法[J]. 生態學報,2013,33(15):4631-4638.
[7] 魚小軍,王 芳,龍瑞軍. 破除種子休眠方法研究進展[J]. 種子,2005,24(7):46-49.
[8] 王 楠,高 靜,黃文靜,等. 赤霉素浸種時長和施用濃度對重度干旱和鹽脅迫下黃芪種子萌發的影響[J]. 生態學雜志,2018,37(8):2339-2344.
[9] He X,Wada Y,Wanders N,et al. Intensification of hydrological drought in California by human water management[J]. Geophysical Research Letters,2017,44(4):1777-1785.
[10] 馮 燕. 苦豆子(Sophora alopecuroides L.)不同種群種子物理休眠釋放的研究[D]. 蘭州:蘭州大學,2011.
[11] Attree S M, Fowke L C. Embryogeny of gymnosperms:advances in synthetic seed technology of conifers[J]. Plant Cell Tissue and Organ Culture,1993,35(1):1-35.
[12] Xia F S,Chen L L,Yan H F,et al. Antioxidant and ultrastructural responses to priming with PEG in aged,ultra-dry oat seed[J]. Seed Science & Technology,2016,44(3):1-13.
[13] 張樹清,張夫道,劉秀梅,等. NaCl對大白菜種子萌發和幼苗生長的影響[J]. 植物營養與肥料學報,2006,12(1):138-141.
[14] 梁 靜,王仲朗,張瑞賓,等. 木芙蓉種子萌發特性研究[J]. 種子,2006(6):41-42,46.
[15] 龔維紅,王月森. 濃硫酸不同處理時間對梧桐種子萌發率及根系生長的影響[J]. 江蘇農業科學,2013,41(3):161-162.
[16] 李振基,宋愛琴. 聚乙二醇(PEG)模擬水分脅迫對木荷種子萌發的影響[J]. 福建林業科技,2007,34(4):1-3.
[17] 于 軍,焦培培. 聚乙二醇(PEG6000)模擬干旱脅迫抑制矮沙冬青
種子的萌發[J]. 基因組學與應用生物學,2016,29(2):355-360.
[18] 施積炎,丁貴杰. 水分脅迫對不同種源馬尾松種子發芽的影響[J]. 山地農業生物學報,2000,19(5):332-337.
[19] 陳士超,王 猛,汪 季,等. 紫花苜蓿種子萌發及幼苗生理特性對PEG6000模擬滲透勢的響應[J]. 應用生態學報,2017,28(9):2923-2931.
[20] 王建科,王 鈞,胡延萍,等. 紫花苜蓿種子萌發過程中對不同鹽脅迫的響應[J]. 分子植物育種,2018,16(1):268-274.
(責任編輯:肖光輝)
