熱穩定性高β-甘露聚糖酶產生菌的篩選、鑒定及酶學性質研究
2020-10-23 11:34:18高偉強陳曉波張春曉
食品工業科技 2020年19期
田 庚,高偉強,陳曉波,張春曉
(河北科技大學生物科學與工程學院,河北石家莊 050018)
β-甘露聚糖酶(β-1,4-D-甘露聚糖甘露聚糖水解酶(β-1,4-D-mannan mannanohydrolases),EC.3.2.l.78)能夠攻擊甘露聚糖骨架的內部β-1,4糖苷鍵,釋放甘露寡糖[1-2]。β-甘露聚糖酶廣泛存在于細菌、真菌、放線菌、植物和動物中[3]。水解產物甘露寡糖(mannan-oligosaccharide,MOS)不僅能夠刺激腸道正常菌群的增殖,抑制病原微生物的生長[1-4],對乳酸菌(Lactobacilli)和雙歧桿菌(Bifidobacteria)生長有益[5],還具有多種生物學活性,如增強動物免疫、調節糖脂代謝、促生長和抗氧化等作用[6-7],MOS還通過調控腸道微生物增強了二甲雙胍的降糖效果[8]。
β-甘露聚糖酶在食品行業應用廣泛,如在面團中添加β-甘露聚糖酶處理15 min的2.0%魔芋葡甘聚糖,能夠顯著抑制二硫鍵的斷裂和二級結構的變化,防止面團的水分散失,維持面團的拉伸能力[9];β-甘露聚糖酶對降低咖啡和果汁的黏度也非常有效[4]。此外,β-甘露聚糖酶作為飼料添加劑應用于養殖業,起到消除抗營養因子的作用[10-11],對提高飼料利用率有重要貢獻;在造紙工業中,與β-木聚糖酶等半纖維素降解酶類協同使用,不僅能夠減少化學漂白劑的用量,減少環境污染,還能提高紙漿白度,改善紙漿性能[12];在紡織工業中能有效去除紡織品所粘附的多余染料,Singh等[13]從KlebsiellapneumoniaeSS11菌株中鑒定了一種熱和堿穩定的β-甘露聚糖酶,用做洗滌劑添加劑,對于去除因甘露聚糖導致的食品染色效果非常好。
β-甘露聚糖酶在工業化應用過程中,部分領域需要高溫處理,如動物飼料制粒過程中需85 ℃處理約3 min[14],在將纖維素發酵生產糖的過程中,利用高溫使原料變得松散[15]。由于大部分β-甘露聚糖酶的最適溫度為40~75 ℃[14],高溫下酶活性大大降低甚至完全失活,因此,篩選熱穩定性高的β-甘露聚糖酶也是目前研究的熱點,如B.subtilis(TBS2)β-甘露聚糖酶在60、70、80、90和100 ℃的半衰期分別為6 h、4.2 h、2 h、20 min和12 min[14];熱和堿穩定的KlebsiellapneumoniaeSS11β-甘露聚糖酶,70 ℃時酶的半衰期為135.91 min[13],B.clausiiS10的β-甘露聚糖酶,其最適溫度為75 ℃,70 ℃孵育3 h,殘余酶活性大于50%,在80 ℃酶的半衰期為0.5 h[16]。盡管這些酶表現出一定的熱穩定性,但在溫度超過80 ℃時,熱穩定性欠佳,限制了其在養殖業及再生能源等領域中的應用,因此開發耐高溫、熱穩定性高的β-甘露聚糖酶具有重要意義。本研究從土壤中分離得到一株降解魔芋膠的地衣芽孢桿菌,并對其產β-甘露聚糖酶的酶學性質進行了研究,為β-甘露聚糖酶的應用奠定了基礎。
1 材料與方法
1.1 材料與儀器
土壤樣品 采自河北科技大學校園內;魔芋膠 湖北惠葡生化科技有限公司;DNS試劑、剛果紅染料、D-甘露糖 北京索萊寶科技有限公司;Trans Taq HiFi酶 北京全式金生物技術有限公司;PCR引物合成及測序 由英濰捷基(上海)貿易有限公司完成;其他試劑 均為國產分析純。
SpectraMaxm i3x多功能酶標儀 美國Molecular Devices公司;My Cycler PCR儀和凝膠成像系統 美國Bio-Rad公司;BIFUGE STRATOS高速冷凍離心機 美國賽默飛世爾科技公司;SW-CJ-2FD超凈工作臺 蘇州泰安空氣技術公司;FA1004N分析天平 上海民橋儀器有限公司;HH-2型數顯恒溫水浴鍋 金壇市海爾電器有限公司;ZWY-2102C振蕩搖床 上海智誠分析儀器制造有限公司;DH6000電熱恒溫培養箱 河北泰斯特儀器有限公司。
1.2 實驗方法
1.2.1 產β-甘露聚糖酶菌的分離及初篩 根據楊苗等[17]的方法并稍作修改,稱5 g土樣于無菌生理鹽水中,在37 ℃、180 r/min搖床上培養2 h;取2 mL土壤懸液加入到富集培養基(魔芋膠5 g/L,蛋白胨5 g/L,KH2PO41 g/L,MgSO40.1 g/L,自然pH)中,37 ℃、180 r/min搖床上培養12 h。富集培養的菌液進行10-5、10-6和10-7稀釋后,取適量菌液涂布于篩選平板培養基(魔芋膠5 g/L,蛋白胨5 g/L,KH2PO41 g/L,MgSO40.1 g/L,瓊脂20 g/L,自然pH)中,37 ℃倒置24 h,每個稀釋梯度設置3個平行。培養24 h后,向平板中加入適量0.1% 剛果紅染液,靜置30 min,觀察水解圈大小,選取水解圈直徑(D)與菌落直徑(d)之比(D/d)較大的菌株,進行復篩,即在篩選培養基上繼續稀釋涂布進行菌種純化,連續2次培養,點種到新的平板上進行剛果紅染色。
1.2.2 菌種的分子鑒定 參照文獻[18]進行基因組DNA提取。以基因組DNA為模版,細菌16S rDNA通用引物16S F1:5′-AGAGTTTGATCCTGGTCAG-3′ 和16S R1:5′-TACGGCTACCTTGTTACGACTTC-3′,使用HiFi Taq進行基因擴增。PCR擴增程序:95 ℃預變性5 min;95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸1 min 40 s,32個循環;72 ℃延伸10 min。
PCR產物測序由英濰捷基(上海)貿易有限公司完成。采用 BLAST軟件對序列進行同源性比較,選取同源性較高菌株的16S rDNA序列,利用MEGA 6.0軟件構建系統發育樹[19-22]。
1.2.3β-甘露聚糖酶酶學性質研究 根據文獻報道的方法稍作修改進行β-甘露糖苷酶的酶學性質研究[23]。
1.2.3.1 粗酶液的制備 將初篩得到的菌株接到種子培養基(蛋白胨10 g/L,酵母粉5 g/L,NaCl 10 g/L,自然pH)中[24],37 ℃、180 r/min培養12 h。按10%的接種量轉接到發酵培養基(魔芋膠5 g/L,蛋白胨5 g/L,KH2PO41 g/L,MgSO40.1 g/L),同樣條件下培養40 h。發酵液12000 r/min離心10 min,收集的上清液即為粗酶液[25]。
1.2.3.2 甘露糖標準曲線的制定 D-甘露糖標準曲線的制定[26]:1 g/L D-甘露糖標準溶液的配制,取六支試管分別加入1 g/L甘露糖標準溶液0、0.06、0.12、0.18、0.24、0.3 mL,依次分別加入ddH2O 0.3、0.24、0.18、0.12、0.06、0 mL。再加入0.6 mL的DNS混勻,沸水浴5 min,冷卻后測定OD540。以D-甘露糖質量(mg)為橫坐標x,以OD540為縱坐標y,繪制標準曲線。每組試驗均設1次空白對照和3次重復試驗,重復試驗取平均值。
1.2.3.3β-甘露聚糖酶活性測定 將魔芋膠浸泡在75%酒精中處理,烘干以除去魔芋膠中含有的還原糖使其變為精魔芋膠,在酶活性測定時作為底物使用[27]。采用DNS法進行酶活性測定,將精魔芋膠溶于pH6.0磷酸鈉緩沖液中配制5 g/L的溶液作為底物,于0.27 mL底物中加入0.03 mL粗酶液,60 ℃水浴10 min,加入DNS試劑0.6 mL,混勻后沸水浴5 min顯色,立即以流動水冷卻至室溫,在540 nm處測定吸光值[28]。
酶活性定義:在一定溫度和pH下,以每分鐘催化底物水解生成1 μmol D-甘露糖所需的酶量定義為一個酶活性單位(U)[29]。
1.2.3.4 pH對酶活性影響 配制pH2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0和12.0緩沖液,與等體積的粗酶液混合,按1.2.3.3的方法測定β-甘露聚糖酶活性,以未經處理的粗酶液作為空白對照。以最適pH的酶活性為100%,計算不同pH的相對酶活性(%),即(特定pH酶活性/最適pH酶活性)×100。
1.2.3.5 酶的pH穩定性 取粗酶液分別在pH2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0和12.0的緩沖液下等體積混合,在37 ℃條件下保溫30 min和2 h。取出酶液使其與0.5%的精魔芋膠底物混合,按1.2.3.3的方法測定剩余β-甘露聚糖酶活性。以最高酶活為100%,分別計算不同pH的相對酶活性(%),即(相同保溫時間特定pH酶活性/該時間最適pH酶活性)×100。
1.2.3.6 溫度對酶活性的影響 將粗酶液進行適當稀釋,加入到0.5%的精魔芋膠底物中,置于不同溫度(30、40、50、55、60、65、70、80、90和100 ℃)下保溫10 min,按1.2.3.3的方法測定β-甘露聚糖酶活性。以最高酶活性為100%,分別計算不同溫度的相對酶活性(%),即(特定溫度酶活性/最適溫度酶活性)×100。
1.2.3.7 酶的熱穩定性 粗酶液分別在溫度60、70、80 ℃下各保溫(2、4、6和8 h),再進行適當稀釋,按1.2.3.3的方法在最適溫度測定β-甘露聚糖酶活性,以最高酶活性為100%,分別計算不同溫度的相對酶活性,即(同一溫度特定時間酶活性/該溫度最大酶活性)×100。
1.2.3.8 金屬離子對酶活性的影響 粗酶液適當稀釋后,分別與10 mmol/L K+、Mn2+、Zn2+、Ca2+、Na+、Cu2+和Mg2+等不同金屬離子溶液等體積混合。在37 ℃條件下保溫1 h,以未加金屬離子的粗酶液作為對照,再按1.2.3.3的方法測定β-甘露聚糖酶活性,以最高酶活為100%,分別計算不同pH的相對酶活性(%),即(特定金屬離子酶活性/未加金屬離子的酶活性)×100。
1.3 數據處理
對酶學性質研究的實驗都進行了3次重復,根據測定數據的平均值和標準誤,采用Origin 2017作圖分析。
2 結果與分析
2.1 菌株的分離初篩
以魔芋膠為唯一碳源,并結合剛果紅染色的方法,從初篩培養基中分離得到D/d比值較大的菌株,純化后,菌落表面濕潤,邊緣不整齊。純化后的菌株點種在篩選培養基上37 ℃培養24 h后進行剛果紅染色,結果如圖1所示,D/d比值平均為1.67。
圖1 分離菌株剛果紅染色結果
2.2 菌種的分子鑒定
菌株經16S rDNA測序,序列提交至NCBI網站(注冊號:MT032348)將序列在NCBI網站上進行BLAST比對,結果表明,該菌株與地衣芽孢桿菌(B.licheniformis)相似性達99.65%。應用Mega6.0軟件構建的系統發育樹,結果如圖2所示,該菌株與B.licheniformisATCC 14580在同一個進化分支上,表明二者親緣關系最近,所以鑒定該菌株為地衣芽孢桿(B.licheniformis)KD-1。
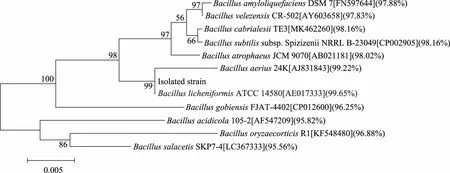
圖2 基于16S rDNA序列構建的系統發育樹
2.3 β-甘露聚糖酶酶學性質
2.3.1 pH對酶活性及穩定性影響 粗酶液與不同pH底物緩沖液等體積混合后,60 ℃保溫10 min,研究pH對酶活性的影響。結果如圖3A所示,β-甘露聚糖酶的最適pH為pH6.0,酶活性達到3.38 U/mL,在pH2.0時保持34%的相對酶活性。在pH3.0~6.0范圍內,隨pH增大,相對酶活性提高;在pH7.0~12.0范圍內,隨著pH增大,相對酶活性降低,但在pH12.0時仍保持49%的相對酶活性。β-甘露聚糖酶在pH5.0~10.0范圍內,保持較高酶活性(大于70%),說明該酶具有較廣的pH耐受范圍(圖3A)。一般來自細菌的β-甘露聚糖酶最適pH在中性范圍[3],如B.licheniformisDSM 13的最適pH在6.0~7.0之間[30],與本研究結果相似;但同是枯草芽孢桿菌屬,其β-甘露聚糖酶最適pH仍存在很大差異,如Bacilussp. N16-5最適pH為9.5[3]。
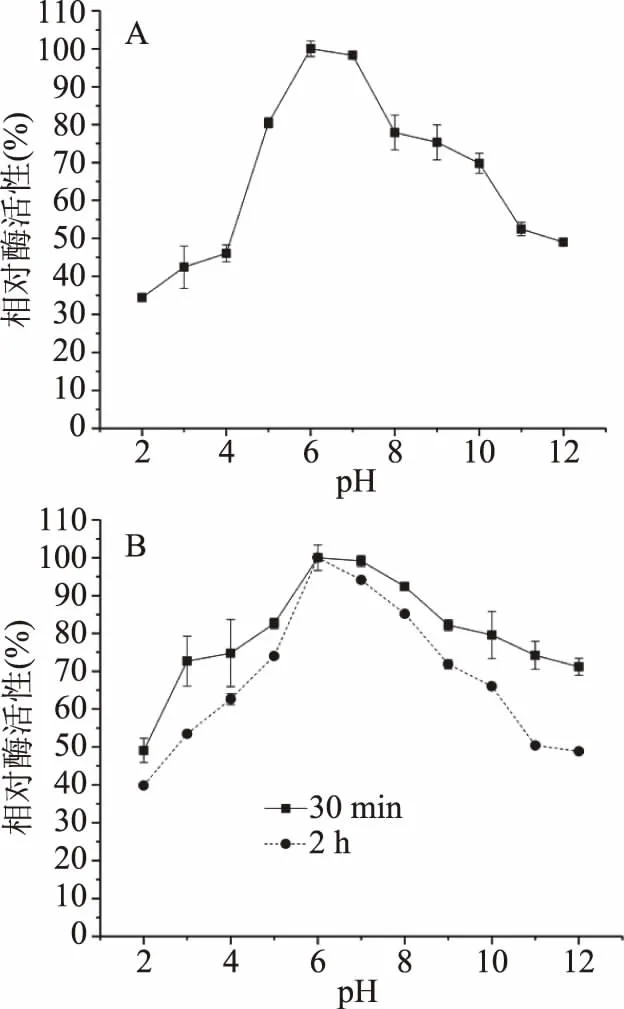
圖3 pH對酶活性及穩定性的影響
酶液與不同pH底物緩沖液等體積混合后在37 ℃孵育30 min和2 h(圖3B),研究pH對酶穩定性的影響。結果表明,該酶在37 ℃孵育30 min后,在pH3.0~12.0之間酶具有較高的穩定性,相對酶活性大于70%,與報道的B.licheniformisDSM 13β-甘露聚糖酶孵育30 min的pH穩定性(pH5.0~12.0)近似,K.pneumoniaeSS11β-甘露聚糖酶在40 ℃孵育30 min,在pH7.0~10.6之間,剩余相對酶活性大于70%[13];孵育2 h后,pH穩定性較30 min有所下降,在pH5.0~9.0之間相對酶活性大于70%,說明該酶在pH5.0~9.0具有較高的pH穩定性。
2.3.2 溫度對酶活性及熱穩定性影響 酶的最適溫度為60 ℃(圖4A),在30~100 ℃范圍內,該酶都保持較高的酶活性,如在30和100 ℃,仍具有高于58%的酶活性,屬于耐高溫酶[31]。60 ℃酶的半衰期(τ1/2)為5.5 h,70和80 ℃酶的半衰期(τ1/2)分別為4.3和4.2 h(圖4B);另外該酶在100 ℃孵育30 min,剩余酶活性仍有31%。以上結果表明該酶的熱穩定性非常高。
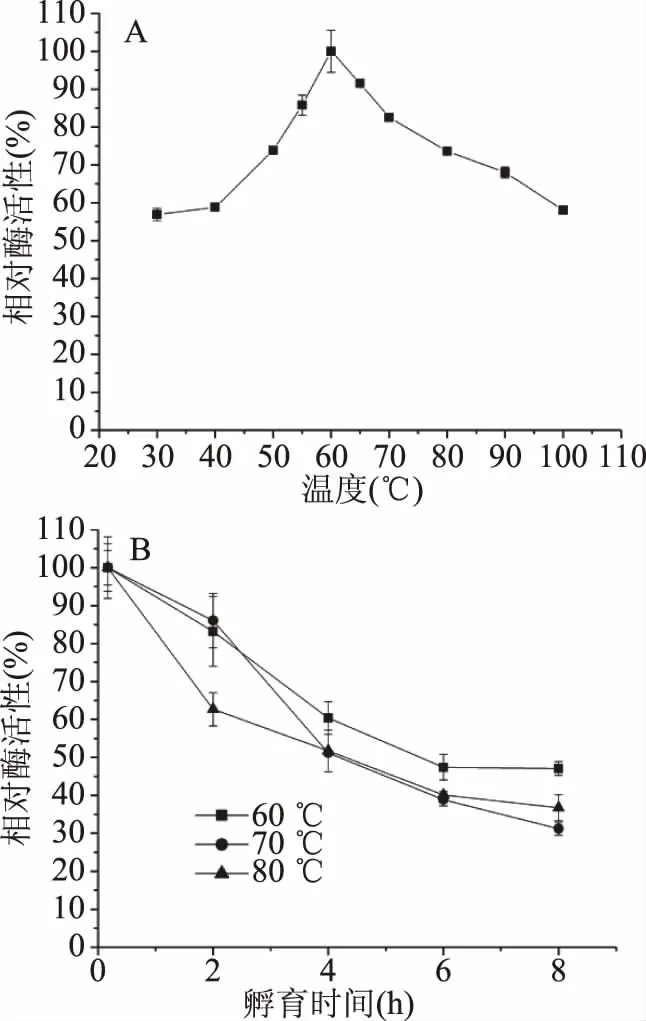
圖4 溫度對酶活性及穩定性影響
本研究β-甘露聚糖酶最適溫度為60 ℃,與B.licheniformisDSM 13[30]、B.subtilisWL-3[32]和B.subtilis(TBS2)[14]等菌株β-甘露聚糖酶的最適溫度接近。盡管這些酶的最適溫度一致,但酶的半衰期有很大不同,如B.licheniformisDSM 13菌株的β-甘露聚糖酶50 ℃酶的半衰期為80 h,60 ℃酶的半衰期迅速下降,僅為3 min,超過70 ℃則酶活性喪失[30],而本研究的B.licheniformisKD-1,利用其發酵液測定的酶80 ℃的半衰期為4.2 h;B.subtilis(TBS2)所產β-甘露聚糖酶80 ℃半衰期為2 h[14],也低于本文報道的β-甘露聚糖酶的熱穩定性,說明B.licheniformisKD-1所產β-甘露聚糖酶是目前報道熱穩定性最高的酶,在飼料制粒中有很大應用潛力。
2.3.3 金屬離子對酶活性的影響 Cu2+和Mg2+具有促進β-甘露聚糖酶活性的作用,分別使酶活性提高了40%和19%(圖5),而其他離子,Mn2+、K+、Zn2+、Ca2+和Na+均對酶活性有不同程度的抑制作用,其中Mn2+對酶活性抑制程度最大,降低了51%的酶活性,Ca2+和Na+僅有微弱的抑制作用,因此在應用過程中或酶制劑制備時適當添加Cu2+或Mg2+有助于提高酶活性。
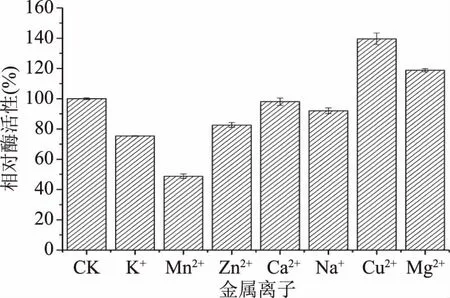
圖5 金屬離子對酶活性的影響
3 結論
本研究分離到一株降解魔芋膠的B.licheniformisKD-1,其產β-甘露聚糖酶的最適pH6.0,最適溫度60 ℃;在pH5.0~9.0和60~80 ℃,酶的穩定性良好,60 ℃酶的半衰期(T1/2)為5.5 h,70和80 ℃酶的半衰期(T1/2)均超過4.2 h,是目前報道熱穩定性最高的一種酶;10 mmol/L的Cu2+和Mg2+明顯促進β-甘露聚糖酶活性,而10 mmol/L的Mn2+明顯抑制酶活性。
