煙草青枯病不同發病階段根際土壤微生物群落變化趨勢分析
2020-11-04 08:16:16黎妍妍彭五星李錫宏
中國煙草科學 2020年5期
關鍵詞:煙草
黎妍妍,王 林,彭五星,李錫宏*
(1.湖北省煙草科學研究院,武漢 430030;2.湖北中煙工業有限責任公司,武漢 430040;3.恩施州煙草公司宣恩縣煙葉分公司,湖北 宣恩 445504)
根際是植物-土壤-微生物相互作用的關鍵微域,每克根際土壤中大約有109個微生物定殖[1]。在自然界中,植物會在其根際土壤中積累病原體,這最終會影響土壤特性[2],造成的根際土壤微生物群落結構的變化又會引起土壤中物質和能量循環、有機質分解與合成等方面的改變[3]。因此,深入認識土傳病害發生過程中根際土壤微生物的變化對于明確根際土壤微生物與土傳病害的互作關系具有重要意義。
煙草青枯病是由青枯雷爾氏菌(Ralstonia solanacearum)引起的一種土傳細菌性病害,每年給煙葉生產造成巨大的產量損失和經濟損失[4]。以往研究通過分析健康煙田和發病煙田之間土壤微生物群落結構差異,探索了微生物與煙草青枯病之間的關系[5-7],但關于青枯病發生過程中根際土壤微生物的變化情況鮮有報道。本研究旨在通過擴增子測序技術,探究煙草青枯病不同發病階段根際土壤微生物群落特征,以期為進一步明確根際土壤微生物群落與青枯病的互作關系提供理論依據。
1 材料與方法
1.1 試驗地點
2017 年在湖北省恩施州宣恩縣椒園鄉涼風村設置3 個煙草青枯病系統調查點(即3 次重復),每個系統調查點面積為132 m2。調查田塊土壤類型為黃棕壤,土壤肥力中等,海拔約900 m,連年種植煙草,煙草青枯病發生嚴重。種植煙草品種為云煙87,移栽期為4 月25 日前后,整個煙草生育期內不施用防治煙草病害的藥物。
1.2 煙草青枯病病情調查
以株為單位,按照國家標準 GB/T 23222—2008《煙草病蟲害分級及調查方法》記載每株煙青枯病發病嚴重度,計算發病率和病情指數。發病率=發病株數/調查總株數×100%;病情指數=。
1.3 煙株根際土壤樣品采集
于烤煙移栽前(0 d),采用5 點取樣法采集各田塊10~25 cm 深的耕層土壤樣品,混合為一個樣品,記作C_0 d。采用同樣的取樣方法,于烤煙移栽后50、75 和100 d 采集煙株根際土壤樣品[與根系結合較緊密的土壤(4 mm 內)],混合后分別記作C_50 d、C_75 d 和C_100 d。每個土樣3 次重復,共計12 個土樣。土樣混勻后,將其置于干冰中帶回實驗室,保存于-80 ℃冰箱中用于DNA 提取。
1.4 土壤微生物群落分析
1.4.1 DNA 提取和PCR 擴增 利用FastDNA Spin Kit 試劑盒(MP Biomedicals,USA)提取土壤總DNA。分別用515F(5′-GTGCCAGCMGCCGCGGTAA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)引物對細菌16S rDNA V4 可變區進行PCR 擴增;用ITS5-1737F(5′-GGAAGTAAAAGTCGTAACAAG G-3′)和ITS2-2043R(5′-GCTGCGTTCTTCATCGA TGC-3′)引物對真菌ITS1 區進行PCR 擴增。擴增體系為30 μL:15 μL Phusion Master Mix Buffer(2×)、3 μL 引物(2 μmol/L)、10 μL DNA(1 ng/μL)模板和2 μL ddH2O。反應程序為:98 ℃ 1 min;98 ℃10 s,50 ℃ 30 s,72 ℃ 30 s(30 個循環);72 ℃5 min(Bio-rad T100 梯度PCR 儀)。PCR 產物經2%濃度的瓊脂糖凝膠電泳檢測合格后,進行文庫構建,使用Illumina Hiseq PE250 測序平臺進行上機測序(諾禾致源生物信息科技有限公司)。
1.4.2 OTU 聚類與物種注釋 利用Uparse 軟件對所有樣品的Effective Tags 進行聚類,以97%的一致性將序列聚類成為OTUs。對OTUs 代表序列進行物種注釋,用Mothur 方法與SILVA 的SSUrRNA數據庫進行細菌物種注釋分析,用QIIME 軟件中的blast 方法與Unit 數據庫進行真菌物種注釋分析,獲得各分類水平上的群落組成。
1.5 數據處理
利用QIIME 軟件計算α 多樣性指數(Sobs、Chao1 豐富度指數和Shannon 多樣性指數)。用SPSS 22.0 分析煙草青枯病發生情況和微生物群落OTU數量、α 多樣性指數、物種相對豐度等在各樣本間的差異(p<0.05 水平)(One-way ANOVA);根據樣品在屬水平的物種注釋及豐度信息,對相對豐度前30 位的細菌屬和真菌屬的相對豐度進行標準化處理,得到標準化值(樣品在該分類上的相對豐度和所有樣品在該分類的平均相對豐度的差除以所有樣品在該分類上的標準差所得到的值,即Z值),然后采用HemI 軟件繪制Heatmap 圖。
2 結果
2.1 煙草青枯病發生情況
對煙草青枯病發生情況進行了調查與分析(圖1)。結果表明,烤煙移栽后100 d 時,煙草青枯病發病率為97.78%,顯著高于移栽后50 d(22.78%)和75 d(29.44%)的發病率;煙草青枯病病情指數為37.41,顯著高于移栽后50 d(3.27)和75 d(5.37)的病情指數。
2.2 土壤微生物群落多樣性分析
對根際土壤細菌和真菌群落α 多樣性指數進行了差異分析(表1)。結果表明,土壤細菌群落多樣性和豐富度整體呈現出逐漸降低的趨勢;C_100 d的土壤細菌群落Sobs、shannon、chao1 指數和OTUs數量分別較C_0 d 降低了17.52%、8.00%、17.89%和16.11%,且Sobs 和shannon 指數均顯著低于其他樣本,OTUs 數量顯著低于C_50 d 和C_0 d。土壤真菌群落Sobs、shannon、chao1 指數和OTUs 數量在各樣本間不存在顯著差異,但C_50 d 樣本的Sobs、chao1 指數和OTUs 數量均低于其他樣本,這3 個指標分別較C_0 d 降低了15.28%、18.25%和15.49%。以上結果表明煙草青枯病發生對土壤細菌群落多樣性和豐富度的影響較大。

圖1 烤煙移栽后不同時期煙草青枯病發生情況Fig.1 The occurrence of tobacco bacterial wilt at different periods post-transplantation
2.3 根際土壤細菌群落結構特征
2.3.1 細菌群落在門水平上的組成 平均相對豐度>1%的細菌門共有10 個(表2),分別為變形菌門(Proteobacteria,35.69%~51.12%)、放線菌門(Actinobacteria,6.69%~18.71%)、酸桿菌門(Acidobacteria,7.81%~18.39%)、芽單胞菌門(Gemmatimonadetes,5.36%~8.67%)、厚壁菌門(Firmicutes,0.76%~5.96%)、擬桿菌門(Bacteroidetes,2.85%~4.94%)、綠彎菌門(Chloroflexi,4.83%~6.72%)、硝化螺旋菌門(Nitrospirae,1.17%~3.94%)、浮霉菌門(Planctomycetes,0.88%~3.53%)和疣微菌門(Verrucomicrobia,1.19%~3.82%)。

表1 土壤微生物群落多樣性和豐富度指數Table1 The diversity and richness indexes of soil bacteria and fungi in rhizosphere soil
在10 個細菌門中,有6 個細菌門的相對豐度在各樣本間存在顯著差異。土壤中Acidobacteria、Nitrospirae 和Verrucomicrobia 的相對豐度隨青枯病病情發展逐漸降低,且C_75 d 和C_100 d 顯著低于C_0 d;與C_0 d 相比,C_75 d 降低幅度分別為47.74%、59.39%和59.42%,C_100 d 降低幅度分別為57.73%、70.30%和68.85%。Proteobacteria 的相對豐度則隨青枯病病情發展逐漸升高,且以C_100 d顯著高于C_0 d、C_50 d 和C_75 d,增加幅度分別為43.23%、35.27%和16.90%。Actinobacteria 的相對豐度以C_75 d 和C_100 d 顯著高于C_0 d,增加幅度分別為179.67%和125.71%。Firmicutes 的相對豐度則以C_50 d 和C_100 d 顯著高于C_0 d。以上結果表明煙草青枯病發生階段(C_50 d、C_75 d 和C_100 d)與烤煙移栽前(C_0 d)土壤細菌群落存在較大差異(表2)。
2.3.2 細菌群落在屬水平上的組成 對土壤樣本中前30 個細菌屬相對豐度的相似性進行了HEMI聚類分析(圖2)。結果表明,C_0 d 與C_50 d 具有相似的細菌群落結構,聚為一類;C_75 d 與C_100 d具有相似的細菌群落結構,聚為一類。30 個細菌屬可以分為4 組(I、II、III 和IV)。烤煙移栽前(C_0 d),土壤中IV 組細菌屬豐度較高,Z值為1.33±0.24,主要包含H16等6 個菌屬;移栽50 d(C_50 d)時,土壤中III 組細菌屬豐度較高,Z值為1.50±0.00,主要包含Blautia等6 個菌屬;移栽75 d(C_75 d)時,土壤中I 組細菌屬豐度較高,Z值為1.34±0.20,主要包含Ralstonia、Massilia、Blastococcus、Gemmatimonas等7 個菌屬;移栽100 d(C_100 d)時,土壤中II 組細菌屬豐度較高,Z值為1.09±0.40,主要包含Sphingomonas、Pseudomonas、Pseudoxanthomonas、Bradyrhizobium、Roseiflexus、Lysobacter和Streptomyces等11 個菌屬。


2.4 根際土壤真菌群落結構特征


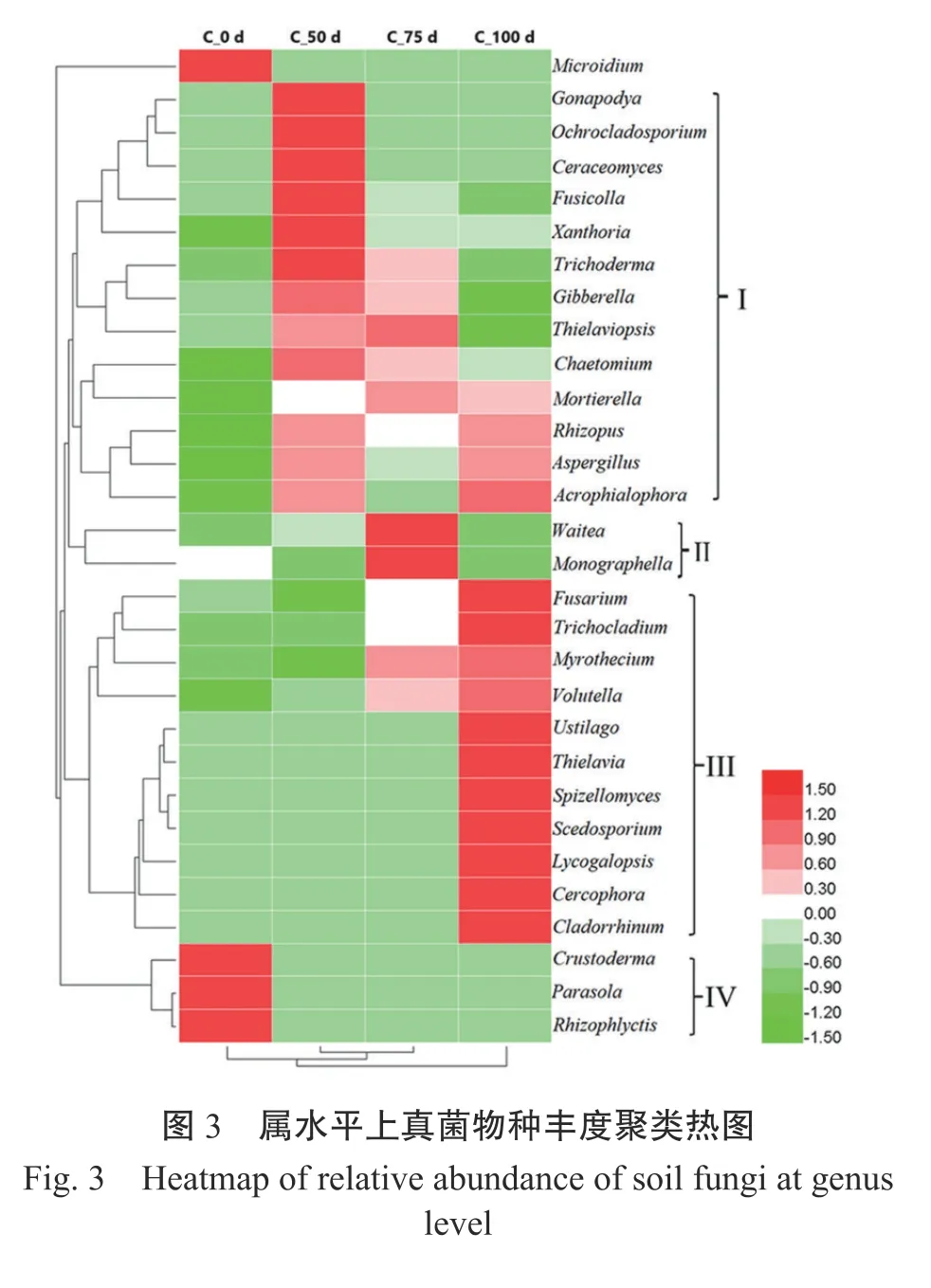

3 討 論
在清江流域煙區,煙草青枯病在病程上可劃分為病害首發期、迅速蔓延期和全面爆發期,分別對應于烤煙移栽后45~67 d、67~97 d 和移栽后97 d至采收結束[8]。本研究運用擴增子測序技術探究了煙草青枯病不同發病階段根際土壤細菌和真菌群落的結構變化,對于明確根際土壤微生物群落與青枯病的互作關系具有重要意義。由微生物多樣性分析結果可知,烤煙移栽75 d 和100 d 時煙株根際土壤細菌群落Sobs、shannon 指數和OTUs 數量顯著降低,這與WEI 等[9]研究結果一致;而真菌群落多樣性指數無顯著變化。以往研究認為,在土壤微生物中,細菌群落多樣性是評價生態系統平衡和生態功能的重要因子[10]。因此,隨著煙草青枯病的流行與蔓延,土壤細菌群落趨向單一化,不利于土壤中微生物種群的平衡[11-12]。
門水平上,根際土壤細菌豐度和結構在烤煙移栽 50 d 和 75 d 發生了明顯的動態變化,Proteobacteria、Actinobacteria 和Firmicutes 的相對豐度顯著升高,Acidobacteria、Nitrospirae 和Verrucomicrobia 的相對豐度顯著降低。與細菌群落相比,門水平上根際土壤真菌群落變化較小,僅Zygomycota 的相對豐度在烤煙移栽75 d 顯著升高。
與移栽前相比,屬水平上根際土壤細菌群落結構于烤煙移栽后75 d 和100 d、真菌群落結構于烤煙移栽后50 d 和75 d 發生明顯變化。從功能上講,根際土壤微生物中包含致病微生物、有益微生物等[13]。致病微生物在根際土壤中定殖,努力突破保護性微生物屏障,克服植物固有的防御機制,從而導致病害發生。通過對屬水平上土壤微生物群落分析發現,致病細菌Ralstonia和致病真菌Fusarium的相對豐度在青枯病發生時明顯升高。本研究中,雷爾氏菌屬Ralstonia僅包含青枯雷爾氏菌R.solanacearum一個種。青枯菌是導致青枯病發生的病原菌,在10種重要的植物病原細菌中,青枯菌位居第2 位[14]。青枯菌相對豐度于烤煙移栽后75 d 達到峰值,這可能與烤煙移栽后,其根系分泌的延胡索酸、肉豆蔻酸、肉桂酸、蘋果酸和草酸等有機酸促進了青枯菌在煙株根部的定殖[15-16]有關;但隨著青枯病的發生,根系活力減弱,分泌有機酸的能力也降低。鐮刀菌屬Fusarium可引起多種植物的萎蔫、根腐[17],其相對豐度隨煙草青枯病病情發展而逐漸升高,于烤煙移栽后100 d 時達到峰值。Fusarium可破壞煙株根部,為青枯菌侵染煙株制造天然的入口[18]。
除了致病微生物外,有益微生物也是根際土壤微生物中重要的一類,且研究較多。它們通過刺激植物根生長、進行根際修復、調控非生物脅迫和控制病害等促進植物生長[13]。在受到病原菌侵染時,植株通常能夠調節根際土壤微生物群落,特異性地招募具有誘導抗病和促進植物生長的有益微生物[19]。本研究結果也證實了這一點。與烤煙移栽前相比,對土傳病原菌具有拮抗作用或對植物生長具有促生作用的Sphingomonas、Gemmatimonas、Pseudomonas、Lysobacter、Streptomyces等有益細菌[20-24]和Trichoderma、Gibberella、Chaetomium等有益真菌[25-27]的相對豐度在青枯病發生階段明顯升高,表明青枯菌在根際和根系的入侵可能與對植物無害甚至有益的某些功能菌具有正相關[28]。
4 結 論
本研究結果表明,青枯病煙田根際土壤細菌群落Sobs、shannon 指數和OTUs 數量于烤煙移栽后75 d 和100 d 顯著降低,真菌群落多樣性指數無顯著變化。與烤煙移栽前相比,屬水平上根際土壤細菌群落結構于烤煙移栽后75 d 和100 d、真菌群落結構于烤煙移栽后50 d 和75 d 發生明顯變化。致病微生物(Ralstonia、Fusarium等)和有益微生物(Sphingomonas、Gemmatimonas、Pseudomonas、Lysobacter、Streptomyces、Trichoderma、Gibberella、Chaetomium等)的相對豐度在青枯病發生階段明顯升高。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
