不同物候期茶樹品種(系)葉綠素熒光特性研究
2020-11-18 08:51:23唐敏翟秀明李解鄔秀宏侯渝嘉
南方農業·上旬 2020年10期
唐敏 翟秀明 李解 鄔秀宏 侯渝嘉
摘? ?要? ?采用PAM-2500便攜式調制葉綠素熒光儀測定不同物候期類型的5個茶樹品種(系)葉綠素熒光參數,分析不同物候期類型茶樹的熒光參數特征,并比較各參數的品種(系)間差異。結果顯示,茶樹對吸收的光能,有79.4%~83.1%通過調節性或非調節性機制以熱量形式耗散掉,20.6%~16.9%用于光化學反應。物候期較早的品種光合中心活性較強,光響應曲線斜率較大,半飽和光強較小,在作用光光強一定時實際光合量子產量高于物候期較晚的品種(系),表現出Y(II)、Alpha、ETRmax等參數顯著高于晚生品種(系),Fo顯著低于晚生品種(系)的熒光特性,且與茶樹物候期的先后具有較高一致性。
關鍵詞? ?茶樹;品種;物候期;葉綠素熒光;光響應曲線
中圖分類號:S184? ?文獻標志碼:A? ? DOI:10.19415/j.cnki.1673-890x.2020.28.003
茶樹是我國重要的經濟作物,其經濟價值與采收時間具有直接的聯系。茶樹光量子利用規律是茶樹光合作用研究的基礎,對早生茶樹資源的鑒定與篩選和種質創新具有重要意義。葉綠素熒光技術是以植物體內光系統Ⅱ(PSⅡ)中葉綠素a的熒光為天然探針,研究和探測植物光合生理狀況的新型植物活體診斷技術,較表觀性的氣體交換指標更具反映內在性的特點[1-3],目前,該技術以其方便、快捷、無損的特點,已經在光、溫度、干旱等逆境生理研究中得到廣泛應用[4-6]。
本研究通過對物候期具有明顯差異的5個茶樹品種(系)的葉綠素熒光參數進行測試分析,旨在探討其物候期與主要熒光參數間的關系及品種(系)間差異,了解不同物候期茶樹品種(系)的光化學啟動特性,為早生茶樹資源的早期鑒定和早生茶樹種質創新提供理論依據。
1 材料與方法
1.1 試驗材料
試驗材料為生長健康,無病蟲害的福鼎大白茶、渝茶3號、渝茶4號、烏牛早和品系0301的5年生成年茶樹,選擇當年生成熟葉片作為測定對象。
1.2 試驗地點
試驗在重慶市農業科學院茶葉研究所試驗茶園進行,地理位置為北緯29°75′、東經105°71′,海拔440 m,屬亞熱帶季風性濕潤氣候。2017—2019年平均年積溫4 787.20 ℃,平均年日照時長809.13 h。
1.3 熒光參數測定
采用PAM-2500便攜式調制葉綠素熒光儀測定熒光參數。測定前,將葉片進行充分暗適應(20 min以上),待葉片暗適應后,照射檢測光,測定初始熒光Fo。然后加飽和脈沖光,測定最大熒光Fm。打開測量光和光化光,每隔10 s照射1次飽和脈沖光,測量光下最大熒光,采用Eilers and Peeters[Ecological Modelling 42(1988) 119-215]的公式擬合光響應曲線。所有參數測定均重復5次取平均值。
1.4 物候期調查
物候期調查所選芽為剪口下第一個芽。各目標品種(系)固定觀察10個芽,每隔1~2 d觀察1次,以樣本數的30%為判斷不同時期的標準,參見《茶樹種質資源描述規范和數據標準》[7]。
1.5 數據處理
數據整理和統計分析采用Microsoft Excel和SPSS Statistics 22進行。
2 結果與分析
2.1 不同茶樹品種的物候期
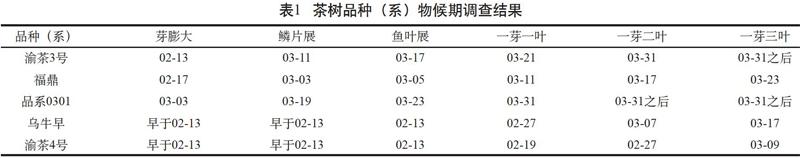
2017—2019年春季,按照《茶樹種質資源描述規范和數據標準》分別對渝茶3號、渝茶4號、福鼎大白茶、烏牛早和品系0301等5個品種(系)在重慶永川地區的物候期進行了調查,調查結果列于表1。
結果顯示,5個品種(系)按達到一芽一葉期的先后時間順序為渝茶4號→烏牛早→福鼎→渝茶3號→品系0301,其中烏牛早、渝茶4號一芽一葉開采時間分別早于福鼎大白茶12 d、20 d,屬于特早生品種,渝茶3號、品系0301分別晚于福鼎10 d、20 d,屬于中生品種和晚生品種。
2.2 不同物候期茶樹品種(系)葉片Fo、Fm、Fv和Fv/Fm的差異
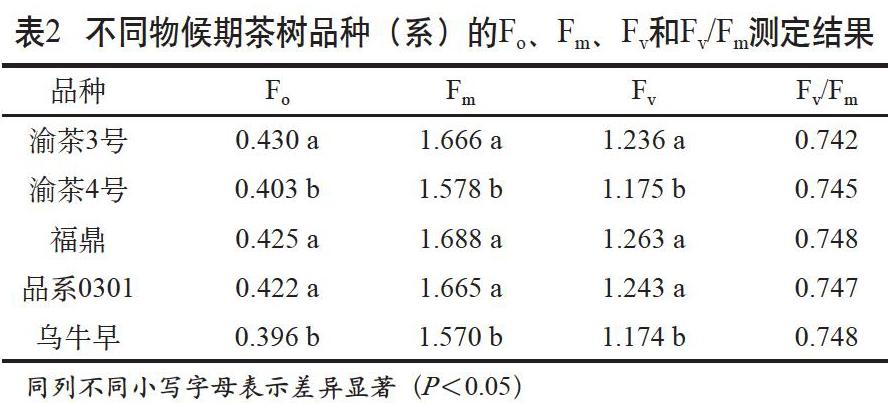
Fm是暗適應后葉片光合機構全部PSII中心都關閉時的熒光強度,這時熒光產量最大,反映了葉片光合中心的全閉合狀態。Fo是暗適應后光合機構全部PSII中心都開放時的熒光強度,來自于天線葉綠素Chla,反映了葉片對光的全開放狀態,與光合中心活性有關[8-9]。其差值Fv反映了葉片光合中心進行光化學反應的“庫”或“能力范圍”[10]。Fv/Fm反映的是植物對其生境長期適應的機制,正常生長植物的Fv/Fm相對穩定。當環境變化對植物造成脅迫或傷害后,Fv/Fm會隨之降低[11-12]。不同物候期茶樹品種(系)的Fo、Fm、Fv和Fv/Fm測定結果見表2。
結果顯示,所試5個品種(系)Fv/Fm差異不顯著,說明參試材料生長正常或受環境影響程度一致。Fo、Fm和Fv因品種(系)不同而表現出明顯差異,其中渝茶4號和烏牛早的Fo、Fm、Fv顯著低于其他3個參試品種(系),表明這2個品種光合中心活性高,但其光合中心進行光化學反應的“庫”較小。
2.3 不同物候期茶樹品種的能量分配差異
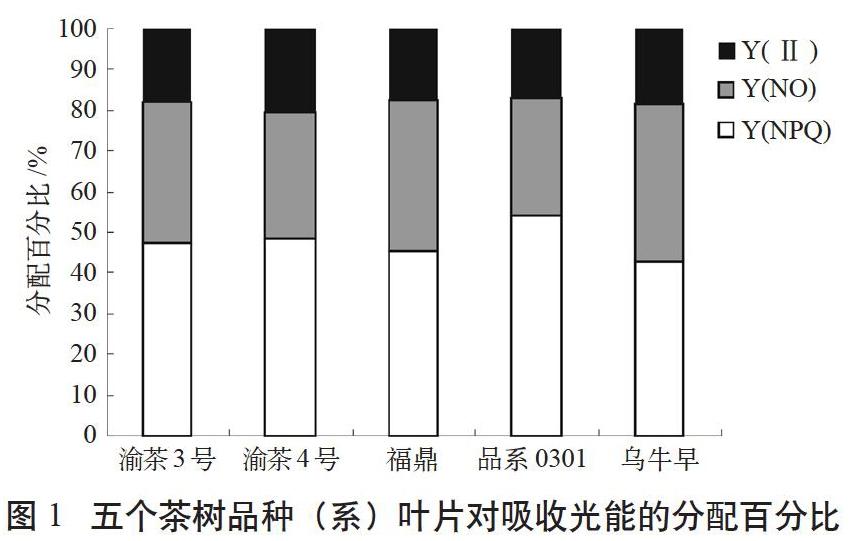
由于葉片接收的光能主要分為光化學能、熱耗散和熒光3個部分被轉化或耗散。這三者之間具有“光化學能+熱耗散+熒光產量=1”的關系,即Y(II)+Y(NO)+Y(NPQ)=1。據此對5個品種的光能利用和分配進行了測定和分析。分別測定了以上5個品種的調節性能量耗散的量子產額Y(NPQ)、非調節性能量耗散的量子產額Y(NO)、實際光量子產量Y(II)(見圖1)。
Y(NPQ)是PSII的調節性能量耗散的量子產量。若Y(NPQ)較高,一方面表明植物接受的光強過剩,另一方面則說明植物仍可通過調節過剩光能,將其轉化為熱而耗散,從而保護自身[13]。Y(NO)是光保護的重要指標,是PSII的非調節性能量耗散的量子產量。Y(NO)較高時,表明光化學能量轉換和保護性的調節機制不足以將植物吸收的光能完全消耗掉[14],說明入射光強超過了植物能接受的程度,這時,植物可能已經受到損傷,或者將要受到損傷。
由圖1可知,所示5個品種(系)茶樹對吸收的光能,有79.4%~83.1%通過調節性或非調節性機制以熱量形式耗散掉,20.6%~16.9%的能量用于光化學反應,推動自身的碳氮循環和生理活動,但不同品種對能量的分配比例各有不同。渝茶4號Y(II)最高,品系0301和福鼎最低,且與渝茶4號差異顯著,烏牛早、渝茶3號居中,為過渡類型,與Y(II)最高和最低的品種均未達到統計學上的顯著差異水平。所試5個品種(系)光合量子產量由大到小的順序為渝茶4號>烏牛早>渝茶3號>福鼎>品系0301,這與一芽一葉物候期觀察結果基本一致。
2.4 不同物候期茶樹品種的光響應曲線
通過擬合光響應曲線(RLC)能反應不同光合有效輻射下葉片通過PSII的電子傳遞速率。本試驗連續測定了葉片在0、13、30、55、87、127、178、235、303、390、503和640 μmol·m-2·s-1共12個光合有效輻射下葉片的電子傳遞速率,并采用Eilers and Peeters(1988)的公式ETR=PAR/(a·PAR2+b·PAR+c),對光響應曲線進行擬合,測定及擬合結果見圖2。
結果顯示,不同物候期茶樹品種(系)在同樣環境條件下,各品種(系)通過PSII的電子傳遞速率(ETR)隨著光強的增加而不斷增加,當光強增大到400~600μmol·m-2·s-1時,ETR增速減緩直至不再增加。所試5 個茶樹品種(系)中渝茶4號的電子傳遞能力處于較高水平,其余品種(系)相對較弱。據報道,ETR與植物凈光合速率呈顯著相關,光合速率與ETR的動態變化一致[15]。這說明渝茶4號的光合能力相對較強。
2.5? ?不同物候期茶樹品種的光啟動速率差異
通過分析光響應曲線的斜率、最大電子傳遞速率和半飽和光強可進一步反應不同物候期茶樹品種間光啟動速率的差異。采用公式(1)(2)(3)對曲線的斜率(Alpha)、最大電子傳遞速率(ETRmax)和半飽和光強(Ik)進行測算。
計算結果列于表3。
結果顯示,烏牛早和渝茶4號光響應曲線斜率最大且半飽和光強最小,說明這兩個品種光化學啟動較快,且在弱光照條件下就可以啟動光化學過程,與這兩個品種物候期較早的田間觀察結果相吻合。隨光照強度增加,烏牛早和渝茶4號通過PSII的電子傳遞速率迅速上升,直至達到最大電子傳遞速率時ETR增速減緩直至不再增加。其中ETRmax以福鼎和渝茶4號最大,烏牛早最小,這間接反映了烏牛早較其他參試品種(系)茶樹雖能較快啟動光化學反應,但也可能更易受到強光脅迫。
3 小結與討論
葉綠素熒光技術作為植物光合生理的一種方便、快捷的無損檢測技術,能準確地反映植物對光能的吸收、利用、分配和耗散過程,從而間接反映不同類型或不同環境下植物的光化學反應速率和最低光能需求的差異。通過分析和比較不同物候期類型茶樹品種(系)的葉綠素熒光參數的數量特征及其種間差異可以看出,茶樹對吸收的光能,總體上有79.4%~83.1%通過調節性或非調節性機制以熱量形式耗散掉,20.6%~16.9%的能量用于光化學反應,但不同物候期類型的茶樹的熒光參數存在較大差異,其中物候期較早的品種(系)光合中心活性較強,光響應曲線斜率較大,半飽和光強較小,在作用光光強一定時實際光合量子產量高于物候期較晚的品種(系),表現出早生品種(系)Y(II)、Alpha、ETRmax參數顯著高于晚生品種(系),Fo參數顯著低于晚生品種(系)的熒光特性,且與茶樹物候期的先后具有較高一致性。進一步研究和利用這些特性與差異,對構建以葉綠素熒光技術為基礎的茶樹早生資源鑒定體系和推動茶樹種質創新具有重要意義。
參考文獻:
[1] Krause GH, Weis F. Chlorophyll fluorescence and photosynthesis: The Basics[J]. Annual Review of Plant Physiology, 1991(42): 313-349.
[2] 王碧瑩,盛連喜,郭靜,等.不同施氮處理下短梗五加與花楸葉綠素熒光參數日變化的比較研究[J].東北師大學報(自然科學版),2019,52(3):107-114.
[3] 李曉,馮偉,曾曉春,等.葉綠素熒光分析技術及應用進展[J].西北植物學報,2006,26(10):2186-2196.
[4] Lamont BB, Lamont HC. Utilizable water in leaves of 8 arid species as derived from pressure-volume urves and chlorophyll fluorescence[J]. Physiol Plant, 2000, 110(1): 64-71.
[5] Lu Congming, Zhang Jianhua. Photosynthetic CO2 assimilation, chlorophyll fluorescence and photoinhibition as affected by nitrogen deficiency in maize plants[J]. Plant Science, 2000, 151(2): 135-143.
[6] Ogren E, Evans JR. Photoinhibitionin situin six species of Eucalyptus[J]. Australian Journal of Plant Physiology, 1992, 19: 223-232.
[7] 陳亮,楊亞軍,虞富蓮,等.茶樹種質資源描述規范和數據標準[M].北京:中國農業出版社,2005.
[8] 許大全.光合作用效率[M].上海科學技術出版社,2002:32-33.
[9] 張紅娜,蘇鉆賢,陳厚彬,等.荔枝成花期誘導性低溫對其葉片光合特性的影響[J].廣東農業科學,2016,10:59-63.
[10] 王蘭蘭,李琦,宋曉卉,等.環境條件對植物葉綠素熒光參數影響研究進展[J].沈陽師范大學學報(自然科學版),2019(4):362-367.
[11] 張守仁.葉綠素熒光動力學參數的意義及討論[J].植物學通報,1999,16(4):444-448.
[12] 許大全,張玉忠,張榮銑.植物光合作用的光抑制[J].植物生理學通訊,1992,28(4):237-243.
[13] 陳景玲,王靜,王謙,等.基于葉綠素熒光的荊條灌叢對栓皮櫟幼苗庇蔭效應研究[J].西北林學院學報,2014(4):46-53.
[14] 劉愛榮,陳雙臣,王淼博,等.高溫脅迫對番茄幼苗光合作用和葉綠素熒光參數的影響[J].西北農業學報,2010(5):145-148.
[15] 顏廷武.不同種源美國紅楓苗期光合特性研究[J].遼寧林業科技,2014(6):24-26.
(責任編輯:丁志祥)
