急性低氧脅迫和復氧恢復對青田田魚幼魚氧化應激和能量代謝的影響
2020-11-27 02:05:36侯懿玲苗藝馨莊淑婷曲憲成劉其根
淡水漁業 2020年6期
關鍵詞:氧化應激
齊 明,侯懿玲,劉 韜,苗藝馨,莊淑婷,曲憲成,劉其根
(1.上海海洋大學,水產科學國家級實驗教學示范中心,上海 201306;2.上海海洋大學,農業農村部淡水水產種質資源重點實驗室,上海 201306;3.上海海洋大學,上海水產養殖工程技術研究中心,上海 201306)
溶氧(DO)是影響魚類生存的重要環境因子之一,生境中溶氧的含量影響其代謝、生長和繁育等生理活動[1]。水體中急性低氧一直是水產養殖中棘手的問題,除輔助人工增氧,更多的是依靠魚類的生理代謝進行適應性調節來緩解[2]。魚類在急性低氧生境中,易誘發機體的氧化應激反應,尤其在低氧后的復氧過程中,氧化應激會更加劇烈[3]。氧化應激會產生過量的活性氧(ROS),擾亂機體內ROS的動態平衡,損傷細胞內的大分子物質并引起代謝紊亂;同時生境中低氧也會造成魚體組織的缺氧,干擾正常的能量代謝[4]。
青田田魚(Cyprinuscarpiovarqingtianensis)又稱甌江彩鯉、田魚,隸屬于鯉形目(Cypriniformes)鯉科(Cyprinidae)鯉屬(Cyprinus),是浙江青田、永嘉一帶一種特有的稻田養殖鯉[5]。稻田生境有別于傳統水產養殖環境,其顯著特征為DO晝夜變化幅度較大并存在長時間低氧期(6~10 h,DO<4 mg/L),同時存在干涸的風險并且無法有效進行人工輔助增氧[6]。青田田魚是如何適應稻田生境,尚未有報道。本研究以青田田魚為研究對象,探討其在低氧脅迫和復氧過程中肝胰臟中氧化應激、呼吸和能量代謝指標的變化,了解青田田魚對溶氧變化的適應性調節機制,為青田田魚對稻田水環境適應性提供佐證,也為其他魚類在急性溶氧變化的生理調控機制研究提供參考。
1 材料與方法
1.1 實驗材料
試驗用魚購自浙江省青田縣愚公生態農業有限公司同一批次繁育的幼魚,選取活力好,健康無損傷,體長(17.12±0.59)cm和體質量(84.55±7.25)g的幼魚運送至上海海洋大學生態與漁業實驗室進行2周暫養。
1.2 試驗設計
暫養時將63尾幼魚隨機平均分配在3個200 L實驗桶(養殖用水體積150 L)中,每日投喂鯉人工配合餌料1次(寧波天邦食品股份有限公司),每日8∶00和18∶00分別使用曝氣自來水換水一次,每次換水量約為養殖水體的1/2。暫養期間水中溶氧為(7.45±0.16)mg/L,水溫為(25.91±1.27)℃,氨氮為(0.19±0.02)mg/L。正式試驗前一天停止投食,但每日正常換水兩次。低氧脅迫實驗前,完成常氧組的取樣(每個實驗桶取3尾,同一實驗桶中3尾魚相同組織混樣),此時的溶氧含量為(7.13±0.28)mg/L,水溫為(25.17±0.41)℃。隨后往水中注入氮氣,待水中溶氧下降到0.5 mg/L左右后,調節氮氣和空氣的注入流量來控制水中的溶氧,分別在第2,4和6 h時進行幼魚樣本取樣,每桶取3尾,脅迫期間水中的溶氧含量為(0.46±0.19)mg/L,水溫為(25.37±0.95)℃。待低氧脅迫試驗取樣結束后,停止氮氣注入并增大空氣注入量,待水中溶氧含量升至7 mg/L時,調節空氣注入量維持水中的溶氧含量,分別在第2,4 和6 h時進行幼魚復氧恢復試驗樣本取樣,每桶取3尾,恢復期間水中的溶氧含量為(7.75±0.17)mg/L,水溫為(25.21±0.87)℃。整個試驗期間,每10 min使用多功能溶氧儀(YSIPro20,USA)監測一次水中的溶氧變化。
1.3 樣品采集與制備
將各時間點每個試驗組的9尾幼魚置于MS-222的水溶液(200 mg/L)中快速麻醉后,用濾紙擦凈幼魚體表水分后尾部靜脈取血1.5 mL。將抽取的血液保存在含有0.05mL肝素鈉溶液(濃度為1.5%)的2 mL離心管中,4 ℃靜置保存 6 h后離心(4 ℃,3 000 r/min,10 min)取血漿,收集的血漿保存于-20 ℃用于血液指標的測定。
取血后的幼魚置于冰板上進行解剖,取下肝胰臟,用濾紙吸干表面血液后置于2 mL離心管中,經過液氮預冷后放置在-80 ℃冰箱保存。后期肝胰臟組織在液氮條件下研磨成粉,置于離心管中進行準確稱重。根據指標測定需要,往離心管中加入9倍(質量體積比)的0.9%的預冷生理鹽水或5~9倍預冷提取液,渦旋振蕩后離心(4 ℃,3 000 r/min,15 min)取上清液,進行相關生化指標的測定。
1.4 樣品測定
肝胰臟和血液生化指標均使用全波長酶標儀(800TS,BIOTEK,美國)和分光光度計(TU-1900,PERSEE,北京)配合南京建成生物工程研究所生產的試劑盒進行測定。肝胰臟的氧化應激指標包括過氧化氫酶(CAT)、谷胱甘肽過氧化物酶(GSH-PX)、超氧化物歧化酶(SOD)和谷胱甘肽-S-轉移酶(GST)活力,丙二醛(MDA)、總谷胱甘肽(T-GSH)、氧化型谷胱甘肽(GSSG)和還原型谷胱甘肽(GSH)含量。呼吸代謝指標包括己糖激酶(HK)、磷酸果糖激酶(PFK)、丙酮酸激酶(PK)、檸檬酸合酶(CS)和乳酸脫氫酶(LDH)活力。能量代謝指標包括肝胰臟中糖原(Gn)和乳酸(LD)含量,血糖(B-Glu)和血乳酸(B-LD)含量。具體檢測方法詳見試劑盒說明。
1.5 數據處理
利用SPSS22.0統計分析軟件對試驗所得的數據進行單因素方差分析(One-way,ANOVA),運用鄧肯(Duncan)多重比較對均值進行差異顯著性檢驗,顯著水平設為P<0.05。試驗所得數據用平均數±標準差(mean±SD)表示。
2 結果與分析
2.1 急性低氧脅迫和復氧恢復對青田田魚氧化應激指標的影響
急性低氧脅迫和復氧恢復對青田田魚肝胰臟中酶促型氧化應激指標的影響見表1。在低氧脅迫階段,脅迫6 h后SOD活力顯著高于常氧組,脅迫2、4和6 h時GST活力均顯著高于常氧組。在復氧恢復過程中,CAT活力在恢復2 h到4 h顯著降低,但各恢復組與常氧組和脅迫6 h組相比無顯著差異;SOD活力在恢復到6 h顯著降低,恢復6 h時 SOD活力顯著低于脅迫6 h,但與常氧組無顯著差異;GST活力在恢復2 h顯著升高且在各處理組中最高,隨恢復時間的延長其活力顯著降低,其活力在恢復6 h和脅迫6 h時無顯著差異,但GST活力在恢復6 h顯著高于常氧組。
急性低氧脅迫和復氧恢復對青田田魚肝胰臟中非酶促型氧化應激指標的影響見表2。MDA含量隨脅迫時間的延長逐步降低且在6 h時其含量顯著低于常氧組;T-GSH含量和GSH/GSSG比例隨脅迫時間的延長先升高后降低,T-GSH含量在脅迫2 h時顯著高于常氧組而脅迫4 h時顯著低于常氧組,GSH/GSSG比例在脅迫2 h和4 h時顯著高于常氧組而脅迫6 h時顯著低于常氧組。在復氧恢復過程中MDA含量先升后降且在4 h其含量為各處理組中最高,恢復6 h時其含量與脅迫6 h時相比無顯著差異;T-GSH含量隨著恢復時間的延長顯著升高,與脅迫6 h和常氧組相比,恢復6 h時其含量無顯著差異;GSH/GSSG比例隨恢復時間的延長升高,恢復6 h其比例顯著低于脅迫6 h和常氧組。
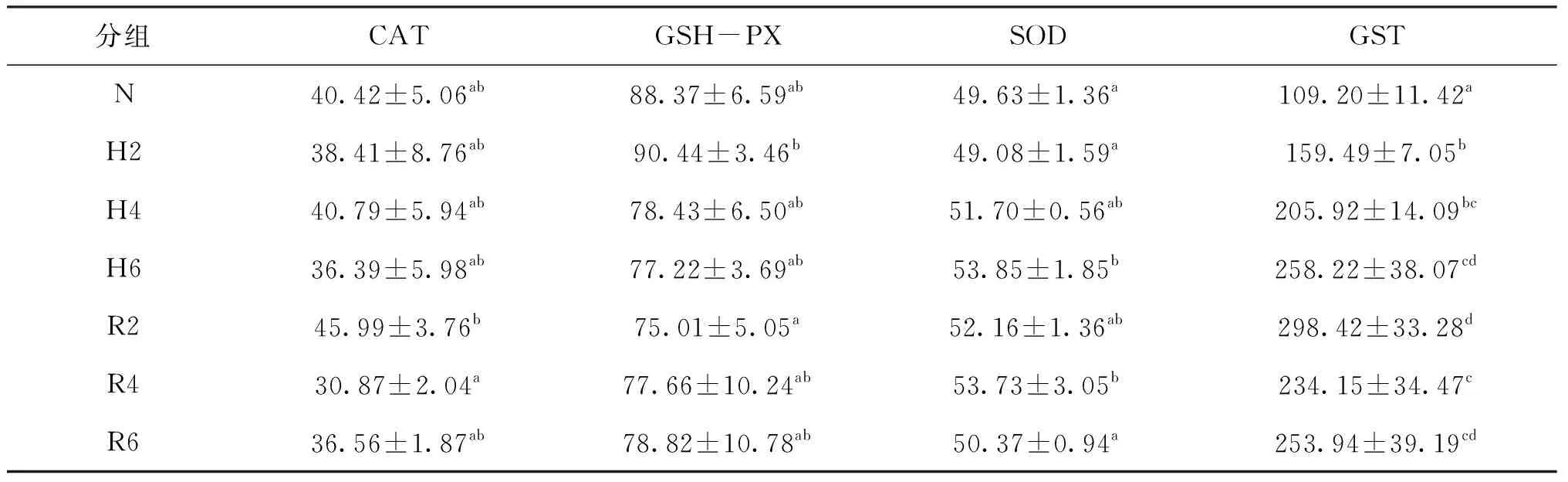
表1 急性低氧脅迫和復氧恢復對青田田魚幼魚肝胰臟中酶促型氧化應激指標的影響Tab.1 Effects of the oxidative index of enzymatic type in the hepatopancreas of juvenile C.carpio var qingtianensis during acute hypoxia and re-oxygenation U/mg prot
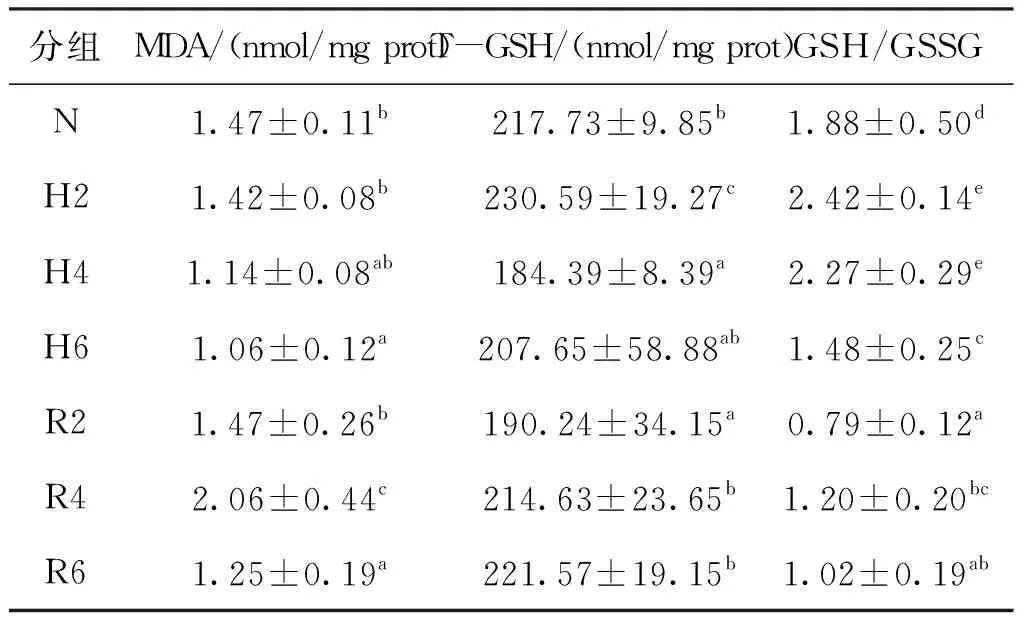
表2 急性低氧脅迫和復氧恢復對青田田魚幼魚肝胰臟中非酶促型氧化應激指標的影響Tab.2 Effects of oxidative index of nonenzymatic type in hepatopancreas of juvenile C.carpio var qingtianensis during acute hypoxia and re-oxygenation
2.2 急性低氧脅迫和復氧恢復對青田田魚肝胰臟中呼吸代謝的影響
由表3可見,脅迫2 h時CS活力顯著高于脅迫6 h;脅迫6 h時LDH活力顯著高于常氧組,其他各呼吸代謝指標在各低氧脅迫處理時間點無顯著差異。經過6 h的恢復,各呼吸代謝指標與脅迫6 h相比無顯著差異,但與常氧組相比,HK活力顯著升高,在恢復2 h時HK、PFK和PK活力為各實驗處理組中最低。
2.3 急性低氧脅迫和復氧恢復對青田田魚的肝胰臟和血液中能量代謝的影響
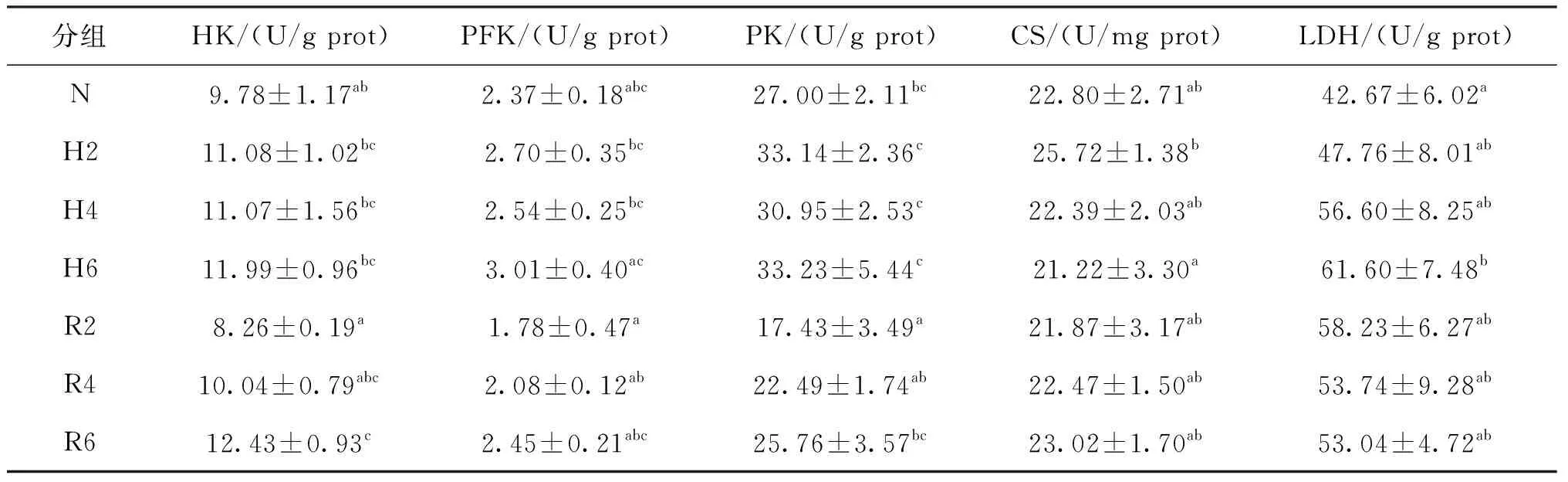
表3 急性低氧脅迫和復氧恢復對青田田魚肝胰臟中呼吸代謝的影響Tab.3 Effects of respiratory metabolism in hepatopancreas of juvenile C.carpio var qingtianensis during acute hypoxia and re-oxygenation
根據表4可知,Gn含量在低氧脅迫過程中顯著降低且各脅迫組顯著低于常氧組;B-Glu含量先降低后升高且脅迫6 h時其含量與常氧組無顯著差異;LD和B-LD含量隨脅迫時間的延長逐步升高且脅迫6 h其含量顯著高于常氧組。
在復氧恢復過程中,Gn、B-Glu、LD和B-LD含量無顯著變化,恢復6 h Gn和B-Glu含量與脅迫6 h時相比無顯著差異;而LD和B-LD含量在恢復6 h時顯著高于常氧組,但與脅迫6 h時無顯著變化。
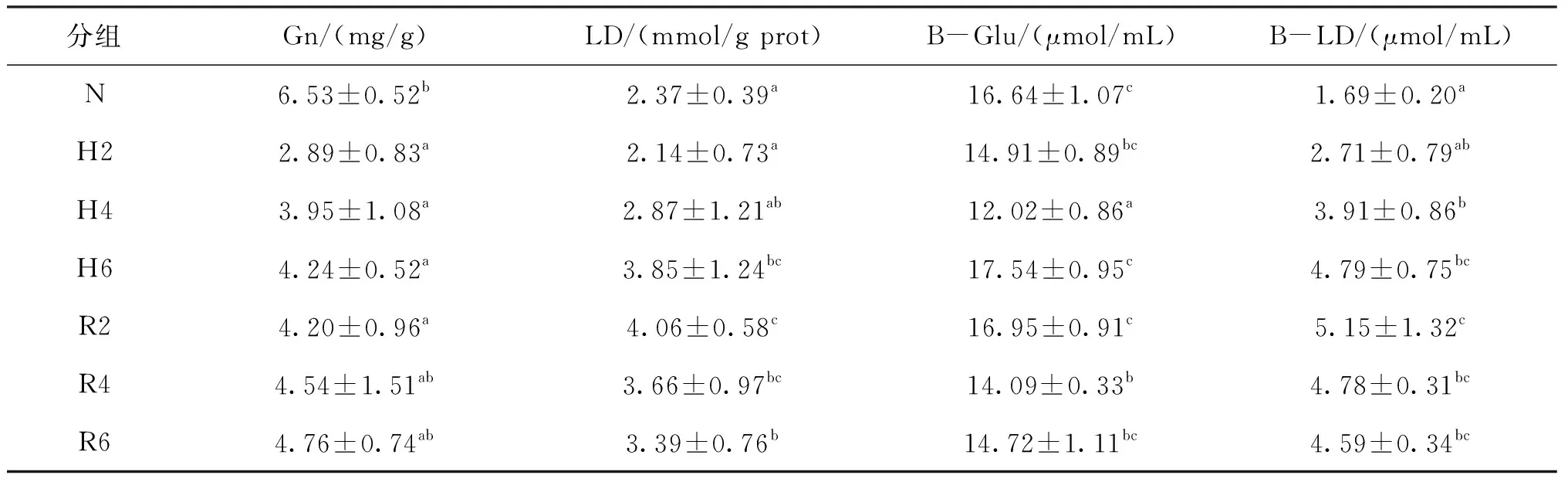
表4 急性低氧脅迫和復氧恢復對青田田魚的肝胰臟和血液中能量代謝的影響Tab.4 Effects of energy metabolism in hepatopancreas and blood of juvenile C.carpio var qingtianensis during acute hypoxia and reoxygenation
3 討論
3.1 急性低氧脅迫和復氧恢復對青田田魚氧化應激指標的影響
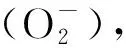
青田田魚肝胰臟中MDA含量在低氧脅迫過程中持續減少,花鱸受到低氧脅迫時也有類似現象,有學者認為ROS的主要來源之一為線粒體內膜的呼吸鏈底物端,氧化應激產生了過量的ROS抑制了脂質氧化酶的活性,隨著低氧時間的延長造成組織功能性缺氧,限制了線粒體電子傳遞鏈給O2過程,減少了ROS的產生,脂質氧化水平得到緩解[9,14,17]。肝胰臟中T-GSH含量和GSH/GSSG比例在這一過程都先升后降,不同的是在低氧脅迫2 h時T-GSH含量顯著高于其他各時間點,但GSH/GSSG比例變化更加顯著,表明肝胰臟通過合成和轉化產生大量的GSH來清除ROS,后期GSH含量的減少也印證了ROS在低氧末期的產生受到抑制[18]。
恢復初期,CAT、SOD和GST活力顯著升高,MDA含量也在此時顯著升高,而T-GSH含量和GSH/GSSG比例顯著降低,這是由于青田田魚再次增強抗氧化防御機制來應對由復氧產生新一輪的氧化應激,這與花鱸[14]和鯔(Mugilcephalus)[19]的結果十分相似,但是不同的是花鱸肝胰臟中SOD、CAT活力并未顯著變化,這可能是由于實驗設計中低氧濃度和復氧恢復時間有所差異。恢復6 h時,除GST顯著高于對照組外,其余氧化應激指標均恢復到常氧組水平,說明青田田魚肝胰臟組織中基本恢復了穩態。
3.2 急性低氧脅迫和復氧恢復對青田田魚呼吸代謝的影響
魚類在低氧壓力下會更加依賴碳水化合物代謝供能,同時低氧使魚類無法通過呼吸獲得正常需氧量,造成局部組織的缺氧,進而改變呼吸代謝方式[20]。糖原是魚體葡萄糖的主要存儲形式,相較于肌糖原,肝Gn的葡萄糖單位含量更高,有研究表明糖原與魚類的耐低氧能力有關[21]。LD既是無氧呼吸產物,也可作為供能代謝的底物[22,23]。HK、PFK、PK、CS和LDH是葡萄糖代謝過程中重要的調控酶,其活力的變化直觀反映組織細胞的呼吸代謝方式[24,25]。同時LD含量和LDH活力也常用來評估魚類的低氧耐受能力[26]。
本試驗中低氧脅迫初期肝胰臟中Gn含量急劇下降,LD含量略微下降,表明Gn分解成葡萄糖,一部分進行肝臟供能代謝,另一部分經過葡萄糖轉運蛋白輸出補充血糖濃度,保障整個機體其他組織的供能代謝[14,27]。HK、PFK、PK、CS和LDH活力均升高,表明青田田魚在感受到外界溶氧變化后,增強能量代謝進行適應性調整,這與瓦氏黃顙魚(Pelteobagrusvachelli)[28]和雜交黃顙魚(P.vachelii♂×P.fulvidraco♀)[26]的研究結果相類似。LD含量無顯著變化,這是由于低氧脅迫初期青田田魚肝胰臟組織以高效的有氧呼吸供能為主。隨著脅迫時間的延長,LD含量和LDH活力顯著升高,HK、PFK、PK活力和Gn含量無顯著變化,而CS活力略微降低,這表明肝胰臟組織缺氧狀況加重,加強了無氧呼吸供能,削弱了有氧呼吸代謝[25,29]。
復氧初期,LDH活力會略微降低而CS活力略微回升,同時PFK、HK、PK活力顯著降低,Gn含量無顯著變化,這是由于隨著溶氧的恢復,組織缺氧狀況得到緩解,無氧呼吸開始削弱,有氧呼吸代謝方式加強。低氧脅迫階段肝胰臟中積累大量的LD,可以異生為葡萄糖進行Gn回補和供能代謝,也可以轉化丙酮酸進入TCA循環[23]。隨著恢復時間的延長,Gn含量和HK、CS活力略微升高,而LDH活力和LD含量降低,這是由于組織缺氧狀況得到進一步的緩解,青田田魚機體有氧呼吸作用得到加強,而無氧呼吸作用相對減弱。復氧末期,LDH、HK活力升高,CS活力略微升高,LD含量逐步降低,Gn含量逐步升高,表明肝胰臟仍然以LD為底物進行糖異生和代謝供能為主,同時加強了糖酵解作用,有氧呼吸開始加強[30]。同時Gn含量恢復到正常水平也說明機體逐步趨于穩態[31]。張國松[28]在瓦氏黃顙魚的研究中,復氧1.5 h其肝胰臟中的CS活力顯著升高而后降低,但在本研究中并未出現這一現象,這可能是青田田魚長期生活在稻田環境中,對急性溶氧變化產生了一定的適應性。
3.3 急性低氧脅迫和復氧恢復對青田田魚血糖和血乳酸的影響
血糖是機體內葡萄糖主要運輸形式,乳酸為葡萄糖厭氧代謝產物,兩者通過血液循環完成組織間能量的再分配[32]。在本試驗中低氧脅迫初中期,血糖含量顯著降低,這是由于低氧帶來的氧化應激壓力,各組織加強對葡萄糖的利用,這與低氧脅迫4 h團頭魴(Megalobramaamblycephala)的實驗結果一致[33]。低氧脅迫后期血糖和乳酸含量顯著升高,這是由于高血糖可以有效地維持機體的能量供給,同時組織缺氧增強了厭氧代謝,造成血乳酸含量的升高,這與黃帶擬鲹(Pseudocaranxdentex)無氧運動后出現高血糖反應和血乳酸含量升高的結果相類似[34]。在復氧過程中,血液中血糖和乳酸含量逐步降低,這是由于在復氧過程中,組織缺氧狀況得到恢復,乳酸異生為葡萄糖進行能量代謝底物的補充,血糖回補臟器組織進行糖原合成和供能代謝,促進機體穩態[23,35]。
綜上所述急性低氧脅迫與復氧對青田田魚生理代謝產生了顯著的影響。在這一過程中青田田魚通過增強氧化應激防御機制,調整有氧和無氧呼吸的供能方式,保障機體能量供給等手段來適應急性溶氧變化。經過6 h復氧恢復后,抗氧化酶活力和糖原基本恢復到常氧組水平,說明青田田魚生理代謝趨于穩態,同時對高含量乳酸的耐受,表明青田田魚有很強的低氧耐受能力和恢復能力。
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
