檫木開花與果實發育規律初步研究
2020-11-27 02:58:52陳宏志姜景民傅愛平
林業科學研究 2020年6期
關鍵詞:質量
陳宏志,劉 軍,姜景民,孫 楊,董 昕,傅愛平
(1. 中國林業科學研究院亞熱帶林業研究所,浙江 杭州 311400;2. 南京林業大學,江蘇 南京 210037;3. 江西省吉安市永豐縣李山林場,江西 吉安 331504)
檫木(Sassafras tzumu(Hemsley)Hemsley)隸屬樟科(Lauraceae)檫木屬(SassafrasJ.Presl),又名檫樹、黃楸木、桐梓樹、鵝腳板,落葉大喬木,自然分布于我國長江流域及其以南地區海拔100~1 900 m的森林中,是山地次生林的主要先鋒樹種和優勢樹種[1-2]。檫木早期生長快,樹冠常居于林冠上層,早春開花,花色明黃,先花后葉,秋季葉色轉紅或黃,色彩醒目,是重要的彩葉景觀樹種之一[3-4];檫木樹干通直,木材花紋明顯美觀,耐腐有香氣,也是優良的用材樹種[5-7],2017年被國家林業局列入《中國主要栽培珍貴樹種參考名錄》。
檫木屬包括3個種,分別為檫木、臺灣檫木(S. randaiense(Hayata) Rehder)和美洲檫木(S.albidum(Nuttall) Nees),為典型的東亞-北美間斷分布屬[2,8],研究表明檫木屬3個種屬于單系發生類群,東亞的2種組成與北美種是相對的姊妹群,并認為臺灣檫木源于大陸檫木[9]。不同于美洲檫木明確的花性別特征,臺灣檫木和檫木在花性別描述上一直存在爭議[10-11]。早期文獻記載檫木為雌雄異株[1],沈卓君等[10]認為這種記載是由于早期樣本采集時出現了錯誤,并總結了前人對檫木花性別的描述,包括單性雌雄異株、兩性或功能上雌雄異株、雜性異株、花兩性以及花近乎兩性等,后續通過對大量花的觀察認為檫木花為兩性花。谷淑芳等[12]和王馨等[13]的研究在一定程度上支持了該觀點,并認為檫木花為雌蕊先熟的兩性花結構。但到目前為止,包括修訂后的《中國植物志》等權威著作尚無對檫木的花性別進行界定[2]。在果實研究方面,僅見沈卓君等[10]與谷淑芳等[12]對果實結構進行過初步觀察,對果實整個發育過程規律尚缺乏系統的研究。為深化對檫木花和果實的生物學認知,本試驗進一步觀察檫木開花與果實發育過程,解析檫木開花與果實發育規律,為檫木種質資源收集評價、保存利用及育種等研究提供參考依據。
1 材料與方法
1.1 試驗材料
取樣點位于浙江省杭州市富陽區城市森林公園(119°57′ E, 30°03′ N),屬亞熱帶季風性濕潤氣候,冬冷夏熱,春季平均氣溫19.1℃,夏季平均氣溫26.5℃,降水充沛。試驗共選取6株無病害且樹齡超過10 a的檫木并編號,于2018年1—7月取樣并觀察。
1.2 試驗方法
自2018年1月起,每周觀察并取樣1~2次,樣品用50% FAA固定48 h后轉入70% FAA中保存備用,其中檫木花的采樣時間為1—3月,果實采樣時間為4—7月,至果實成熟為止。
檫木開花規律:觀察并記錄檫木混合芽芽鱗開裂期、初花期(開花數 ≥ 5%時記為起點)、盛花期(開花數 ≥ 50%時記為起點)及末花期(開花數 ≥ 95%時記為起點)起始時間[14];用高枝剪于盛花期自每株檫木上剪取30個花枝,統計剪取的花枝上每個混合芽包含的花序數、每個花序花數量,測量花梗長和花徑大小。
檫木果實發育規律:自檫木幼果形成開始,每周取樣1次,利用游標卡尺測定果實橫徑與果實縱徑(每重復50個單果),測定果實鮮質量(稱量50個果實總質量后取平均值),105℃殺青30 min后于70℃烘干至恒質量,試驗設置3次重復。計算果形指數、果實含水率和果實干質量比例并繪制果實生長發育曲線,其中:果形指數 = 果實縱徑/果實橫徑;果實含水率 =(果實鮮質量 - 果實干質量)/果實鮮質量;果實干質量比例 = 果實干質量/果實鮮質量。
檫木花與果實結構:樣品采集固定后,利用MDG33型體視顯微鏡(徠卡公司)和石蠟切片觀察檫木花和果實結構。其中石蠟切片制作方法參照王馨等[15]方法,經脫水、透明處理后用石蠟包埋,切片厚度6~8 μm,利用Imager A2型研究級顯微鏡(蔡司公司)觀察并拍照。
采用Excel2003軟件進行數據處理,利用Origin2018軟件作圖。
2 結果與分析
2.1 檫木花結構特征觀察
利用體視顯微鏡和石蠟切片對檫木花整體及組成部位進行觀察,主要結果如下:檫木花由花被片6枚、雄蕊12枚和雌蕊1枚組成(圖1-A)。花被片呈披針形,長約3 mm,分2輪排列,每輪3枚;12枚雄蕊分4輪排列,最外2輪每輪雄蕊3枚,由花藥和花絲組成,長約3 mm;第3輪雄蕊3枚,在其基部著生2枚腺體,腺體成熟時呈杏黃色或橘紅色,可吸引昆蟲授粉,體式顯微鏡下觀察到腺體外部呈馬蹄形(圖1-B),石蠟切片觀察發現其含有大量油性細胞(圖1-C);第4輪雄蕊3枚,外形呈倒三角形,圍繞雌蕊排列,形態退化無育性且較前3輪體積小,長約1.5 mm(圖1-D),試驗中觀察到的花多為該結構。
作者還觀察到另一種檫木花結構類型,其由花被片8枚、雄蕊16枚和雌蕊1枚組成(圖1-E),花被片8枚分2輪排列,16枚雄蕊分4輪排列,具體表現最外2輪雄蕊每輪4枚,第3輪雄蕊4枚,第4輪雄蕊4枚,其中前3輪雄蕊可育,第4輪雄蕊退化;檫木花雌蕊位于整個花的中央,由柱頭、花柱和子房3部分組成,柱頭具有乳頭狀突起(圖1-F),子房上位,包含1室1胚珠(圖1-G);可育雄蕊花藥4室(圖1-H),花藥囊朝向雌蕊,授粉時花粉囊破裂花粉外散(圖1-I),雄配子體經花柱進入胚囊與卵結合并最終完成授粉。
2.2 檫木開花時序特征
檫木開花過程可分為4個階段,分別為I-芽鱗開裂期、Ⅱ-初花期、Ⅲ-盛花期、Ⅳ-末花期,整個過程持續約60 d。檫木成花的芽位于枝條頂端(圖2-A),是由簇生的花序與新枝生長錐組成的混合芽,芽外部覆蓋數目不等的黃褐色芽鱗,其大小與所包含的花序數目有關。
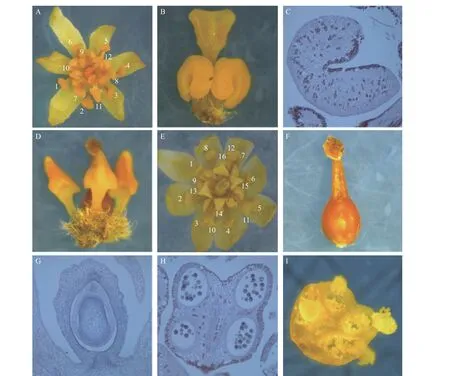
圖 1 檫木花結構特征Fig. 1 The flower structure characteristics of S. tzumu
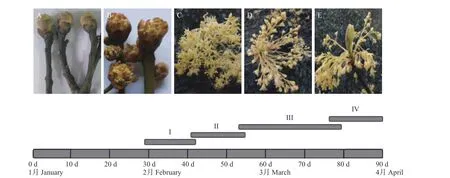
圖 2 檫木開花動態Fig. 2 Floral dynamic of S. tzumu
混合芽于1月底至2月初體積達到最大后芽鱗開裂,簇生狀總狀花序裸露(圖2-B),之后花序不斷伸長,視氣溫條件及發育情況逐漸開花。2月下旬至3月中旬為盛花期(圖2-C),每個混合芽平均可形成花序4.75~9.50個,每個花序平均成花8.25~14.50朵,每個混合芽可成花約68~92朵,且每個混合芽成花量在同株不同混合芽間差異大,平均花徑8.76~11.01 mm,花梗長6.95~7.65 mm(表1)。3月中旬開始授粉,該時期腺體呈杏黃色或橘紅色,并分泌香味物質,花粉囊破裂,花粉外散,吸引昆蟲授粉,完成授粉后花被片閉合、柱頭萎蔫(圖2-D)。3月下旬進入末花期,該時期子房開始膨大,之后花被片及雄蕊逐漸脫落并形成幼果,位于花序中央的新枝生長錐開始生長(圖2-E)。
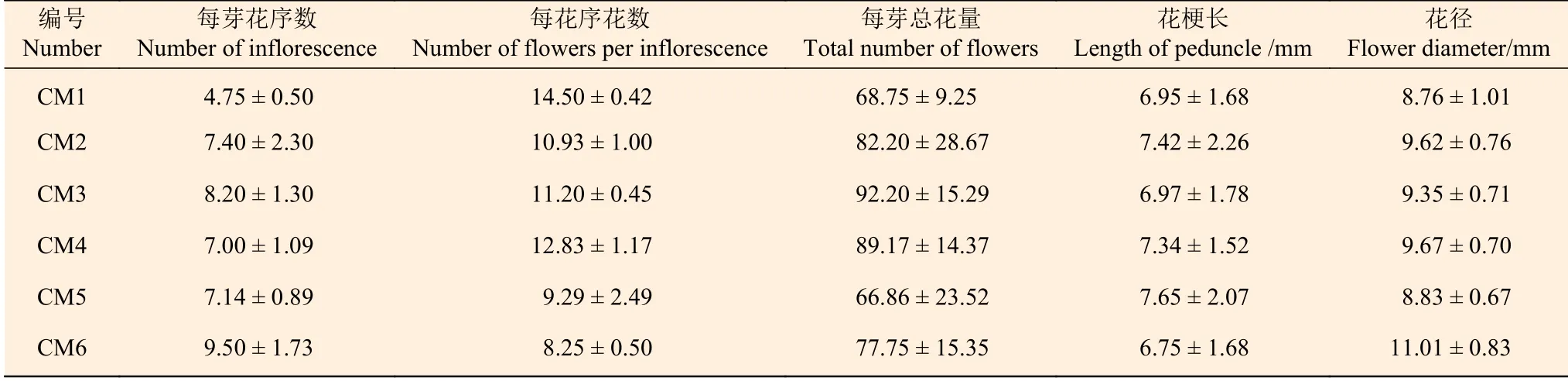
表 1 檫木花部結構數量特征Table 1 The quantitative characteristic of flower structure of S. tzumu
2.3 檫木果實結構變化特征
檫木果實由果皮和種子兩部分組成,按照果實發育特征,可將果實發育分為幼果期、果實膨大期、果實變色期和果實成熟期。檫木花3月中下旬完成授粉后子房膨大,4月初可觀察到幼果,幼果果皮呈綠色,解剖可見內部物質呈白色絮狀,種皮呈白色,果皮厚度約0.22 mm(圖3-A)。5月初解剖新鮮果實(第5周),可觀察到對稱的兩片子葉平伏于胚乳上,兩片子葉中間為胚芽,位于果腔上方(圖3-B),之后子葉吸收胚乳營養不斷生長,體積逐步增大(圖3-C);至6月中旬,胚乳被子葉完全吸收,2片子葉充滿整個果腔(圖3-D),此時種皮已由白色變為黃色,生長1~2周后種皮硬化并由黃色變為黑色,果皮厚度約為0.93 mm,并開始由綠色向黑色轉變,果皮細胞介質逐漸軟化并最終呈漿狀,至果皮成紫黑色時果實完全成熟(圖3-E),整個發育過程持續12周,最終形成漿果狀核果,種子黑色呈圓形,直徑約5.12 mm,為雙子葉無胚乳種子。此外,試驗還觀察到8.53%~27.22%不等的非正常發育果實,該類果實體積與正常果實無差異,具有正常發育的果皮和種皮,但未觀察到種胚(圖3-F)。
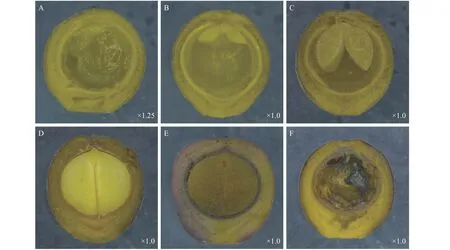
圖 3 不同發育時期檫木果實結構特征Fig. 3 Fruit structure characteristics of S. tzumu at different developmental stages
2.4 檫木果實果形與質量變化特征
在檫木果實發育初期(第1—2周),果實鮮質量和體積緩慢增加,果實縱徑大于果實橫徑,第3—8周果實橫徑與果實縱徑快速增加,8周后果實生長速度降低,果實體積緩慢增加,至第10周果實體積基本達到最大,此時果實橫徑與縱徑分別為6.89 mm和7.14 mm,果形指數1.04(圖4-A),果實外形呈圓形;第3—5周果實鮮質量快速增加,之后生長速度減緩,至第10周果實鮮質量達到最大,單果質量0.17 g,在整個發育過程中果實體積和鮮質量均表現為“慢-快-慢”的變化規律。
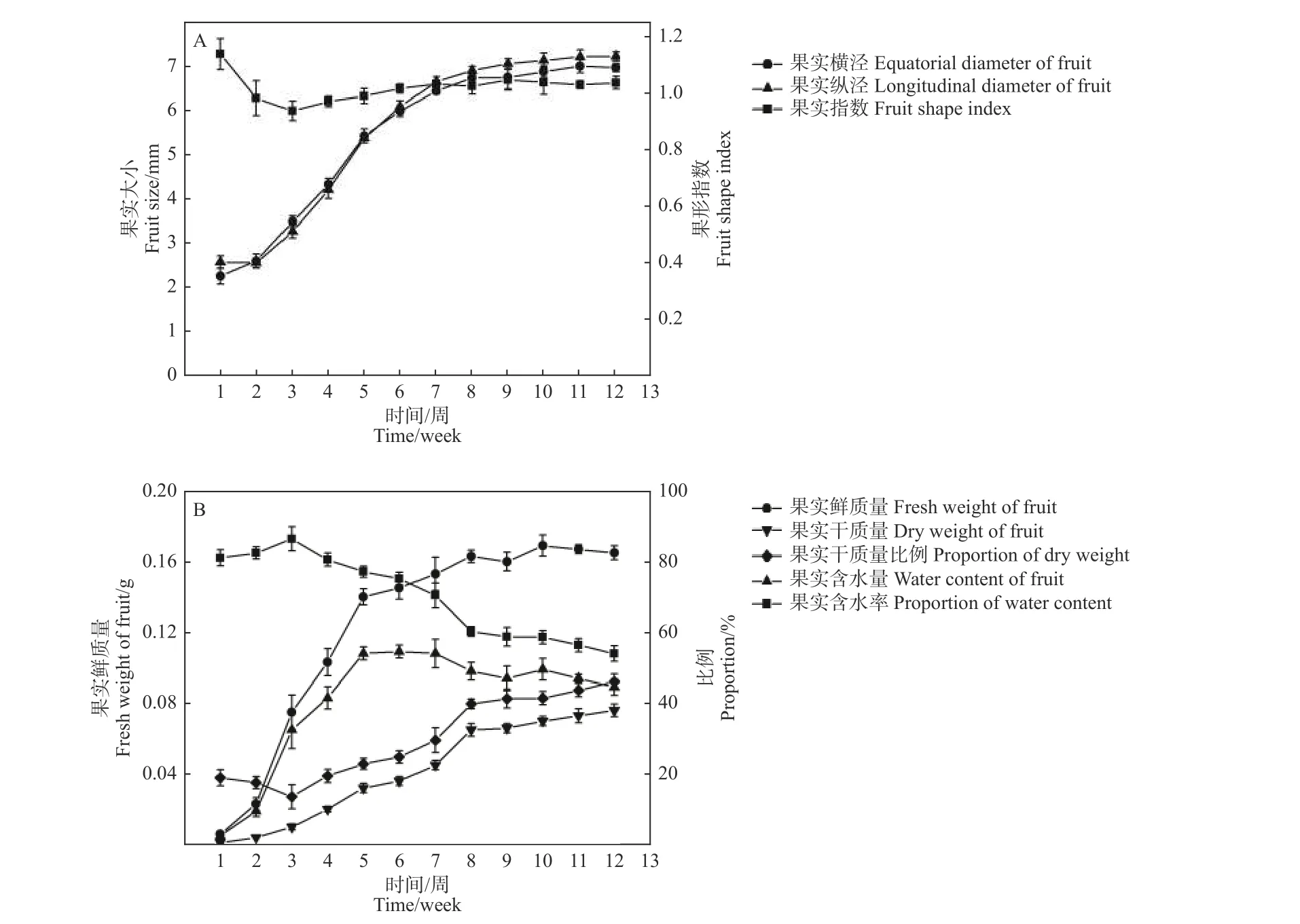
圖 4 檫木果實大小與質量變化規律Fig. 4 Changes of fruit size and weight of S. tzumu
檫木果實發育早期含水率達80%以上(第1—3周),果實干質量低;隨著種胚的形成與生長,果實含水率降低,果實干質量呈線性增加,果實含水量則隨著果實生長先升高后降低的變化特征,其中第5周時含水量最高為0.11 g,占該時期果實鮮質量的77.15%,至果實成熟時含水量降為0.09 g,占果實鮮質量的53.93%,單果干質量0.08 g,占果實鮮質量的46.07%(圖4-B)。以果實發育周期為橫坐標(x),分別以果實鮮質量、果實含水量、果實干質量、果實橫徑和果實縱徑為縱坐標(y)繪制散點圖,數學模型擬合顯示相關指標均與果實發育周期存在一定的函數關系,其R2值均達到0.900以上(表2),表明模型可以較準確地反映檫木果實生長發育的動態變化規律。
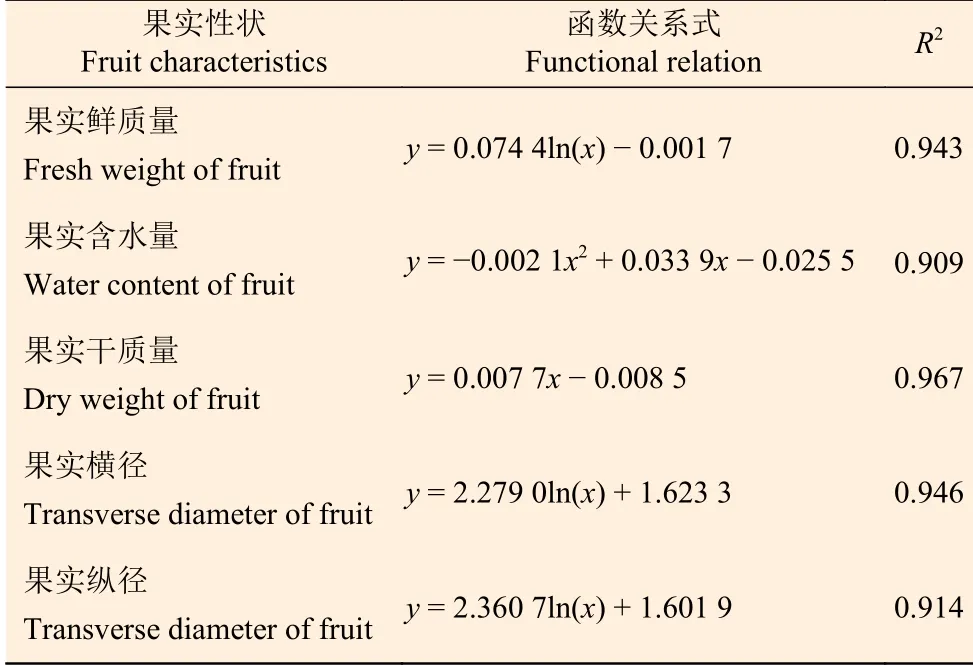
表 2 檫木果實性狀與果實發育周期函數關系Table 2 The relationship between fruit characteristics and develpment periods
3 討論
明確植物開花規律和結構特征是研究植物生活史、了解植物傳粉機制、繁殖系統以及資源收集利用等工作的前提[16-17]。檫木成花的芽1月底2月初芽鱗開裂,群體水平上花期自2月中旬至3月下旬,持續約45 d;在花結構方面,不同于美洲檫木單性花、雄花雄蕊9枚、無退化雄蕊和退化雌蕊、雌花具6枚退化雄蕊的分類特征[18],東亞分布的姐妹種——臺灣檫木和檫木則以花具有正常雌雄蕊結構以及內輪具3枚形態退化雄蕊為主要特征,以至于曾將東亞種作為不同屬或亞屬[2,9]。但關于東亞2個種的花性別長期存在爭議,研究表明臺灣檫木花藥2~4室,花為雌蕊先熟的雌雄異熟兩性結構,應稱為同步或異步雌雄異熟[11,19],檫木具正常兩性花結構,雄蕊花藥4室,雌蕊先于雄蕊成熟[12-13]。調查發現檫木開花量大,每混合芽可成花68~92朵,主要依賴昆蟲授粉,觀察到的花均具有雄蕊和雌蕊結構,表現出兩性花的特征,試驗中未發現單性花,可育雄蕊花藥4室,且供試檫木植株均能正常結實;試驗觀察到2種不同花結構類型,一種由花被片6枚、雄蕊12枚和雌蕊1枚組成,這與多數文獻中所描述的花結構特征一致[2,12],另一種由花被片8枚、雄蕊16枚和雌蕊1枚組成,但該類型花數量較少,關于該類型的花能否正常授粉結實,以及兩種類型的花是否同時存在于同一花序上等問題有待進一步觀察。
檫木果實生長呈現“慢-快-慢”的變化規律,4月初幼果形成,此時新枝生長錐開始生長,與幼果生長形成競爭,造成檫木果實初期生長緩慢[12];4月中下旬檫木葉發育完成并開始提供光合產物,果實進入快速生長期,果實鮮質量快速增加,種胚體積增大,果實含水率降低,果實干質量增加;至6月底進入成熟期,該時期細胞介質成漿汁狀,形成漿果狀核果,果托淺杯狀具長柄,果皮紫黑色被白粉,果核近球形,種子為雙子葉無胚乳種子[12,20]。試驗觀察到檫木果實中存在8.53%~27.22%的無種胚果實,其可能與胚囊僅少數正常發育、雌配子體很少出現數目齊全的胚囊核以及珠心組織存在解離等現象有關[19,21],另一方面,檫木依賴昆蟲傳粉,花雌蕊先于雄蕊熟,且柱頭可授期僅為8 d,由于檫木花中存在花粉敗育現象且花粉活力僅能維持32 h[21],若遇早春嚴寒霜凍或授粉期陰雨天氣等因素,極易造成檫木授粉失敗[10-11],進而導致檫木雖然開花量大,但結實率低甚至不結實現象的產生。
4 結論
檫木開花與果實發育過程觀察結果表明:供試檫木植株群體水平上花期自2月中旬持續至3月下旬,開花過程可分為芽鱗開裂期、初花期、盛花期、末花期4個階段;觀察到少見的花被片8枚、雄蕊16枚及雌蕊1枚的花結構;檫木花具有正常雌雄蕊結構,能夠正常授粉并結果,進一步支持檫木花為兩性花的觀點;檫木果實發育可分為幼果期、果實膨大期、果實變色期以及果實成熟期4個階段,發育過程持續12周,果實為漿果狀核果,種子為雙子葉無胚乳種子。研究結果進一步深化了對檫木生物學特性的認知,并為檫木種質資源收集、保存利用及育種等研究提供了參考依據。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
