滇東南典型常綠闊葉林土壤酶活性的海拔梯度特征
2020-11-27 02:58:54呂晶花任玉連楊羅平王東旭
林業科學研究 2020年6期
李 聰,呂晶花,陸 梅*,任玉連,杜 凡,陶 海,楊羅平,王東旭
(1. 西南林業大學生態與環境學院,云南 昆明 650224;2. 西南林業大學林學院,云南 昆明 650224;3. 貴州大學生命科學學院,貴州 貴陽 550025)
土壤酶是一類主要來源于土壤中動植物體、微生物體的細胞分泌物釋放的生物催化劑[1],它不僅在土壤養分循環及能量代謝方面扮演重要的角色,而且是評價土壤肥力、土壤質量和健康狀況的一個重要生物學指標[2]。土壤酶活性的時空變化受植被類型[3]、土壤生物[4]、土壤水熱條件[5]及土壤養分[6]等生物與非生物因素的共同調控。研究表明,海拔變化會引起區域小氣候、植被群落和土壤理化性質等環境因子的梯度效應,進而顯著影響土壤酶活性[7-8]。任玉連等[9]研究發現,隨著海拔梯度變化,植物群落組成、凋落物性質、微氣候及土壤理化性質等環境因子均發生相應改變。姚蘭等[10]認為,森林植被類型能夠直接或間接地改變土壤活性有機碳/氮組分及含量,從而顯著影響土壤酶活性。因此,土壤酶活性能夠對海拔引起的氣候、植被類型和土壤理化性質等環境因子變化做出響應。然而,調控不同區域土壤酶活性的主導因子可能存在一定差異性,使土壤酶活性對微氣候、植被與土壤沿海拔梯度變化的響應存在地區差異。因此,開展不同地理區域森林土壤酶活性的海拔梯度變化及影響機制研究,不僅有助于理解環境因子變化對森林土壤酶活性及土壤生態學過程的作用機制,而且有助于探明森林土壤對全球變化響應的大小、方向與過程。
云南文山國家級自然保護區地處典型南亞熱帶季風氣候區,森林植被群落結構隨海拔升高變化明顯,且不同海拔植被帶的水熱條件、群落多樣性及土壤理化性質存在顯著的空間異質性,是研究亞熱帶不同海拔梯度森林植被與土壤相互作用的理想場地。因此,本研究選擇文山自然保護區3個不同海拔的典型植被類型(亞熱帶季風常綠闊葉林、半濕潤常綠闊葉林和濕性常綠闊葉林)為研究對象,研究土壤酶活性隨海拔變化的規律及其影響因素,進而闡明土壤酶活性對海拔高度變化的響應特征,為理解該保護區不同海拔“水熱-植被-土壤”之間相互作用對土壤酶活性影響的生態學過程及機制提供理論與數據支撐。
1 研究區概況
文山國家級自然保護區(23°21′00″~23°24′00″ N,104°41′00″~104°41′00″ E)地處滇東南低緯度高原,屬中山山原地貌。該區位于南亞熱帶季風濕潤氣候區,具氣溫高、雨量多及濕度大等特點。保護區地勢起伏平緩,相對高差683 m。年均溫差沿海拔升高由21.1℃降至12.4℃,年均降水量由1 050 mm增至1 630 mm,沿海拔自下而上形成了亞熱帶季風常綠闊葉林、半濕潤常綠闊葉林、濕性常綠闊葉林、苔蘚常綠闊葉林及山頂苔蘚矮林的天然植被垂直分布景觀和森林生態系統[11]。
2 研究方法
2.1 樣地設置
選取文山自然保護區內3個不同海拔的典型植被帶,即亞熱帶季風常綠闊葉林(ME)、半濕潤常綠闊葉林(SE)和濕性常綠闊葉林(HE)。每個植被帶隨機布設3個重復樣地,樣地大小30 m ×30 m(間距約 > 200 m)。樣地基本情況詳見表1。

表 1 樣地基本情況Table 1 The basic situation of the sample plots
2018 年10月,按照上述每個植被類型的3個重復樣地,在每個樣地中采用對角線法選取3個1 m × 1 m取樣點,除去地表覆蓋的凋落物,挖掘寬1 m、深0.8 m的土壤剖面,沿土層自上而下分層采樣(0~10、10~20、20~30、30~40、40~50 cm),剔除石礫、根系等雜物后,用四分法取適量土壤裝入無菌自封袋中,低溫帶回實驗室。將各土壤樣品進行如下處理:約200 g用于土壤含水量測定;約1 kg冷藏于冰箱中,于2周內測定土壤酶活性;約1 kg置于室內自然風干、磨細過篩,用于測定土壤基本性質。各植被帶內3個重復樣方測定的均值作為各植被類型土壤酶活性和理化性質指標。
2.2 土壤理化指標測定
氣溫與降水量數據來源于《云南文山自然保護區綜合科學考察報告》[11]。土壤理化指標測定參照中華人民共和國土壤檢測方法林業行業系列標準[12]。土壤含水量采用烘干法;土壤容重采用環刀法;土壤pH值采用(水土比 1:2.5)電位法;總有機碳采用重鉻酸鉀氧化-外加熱法;全氮采用半微量凱氏定氮法;土壤全磷采用酸溶-鉬銻抗比色法;速效磷采用氟化銨-鹽酸浸提比色法;速效氮采用堿解-擴散法;全鉀采用堿溶-火焰光度計法;速效鉀采用乙酸銨浸提-火焰光度計法。每個土壤樣品至少重復測定3次,取平均值。土壤溫度采用地溫計測定。
2.3 土壤酶活性測定
土壤酶活性測定方法參照《土壤酶及其研究方法》[13]。過氧化氫酶(Cat)采用KMnO4滴定法,以1 g·h-1土壤消耗0.1 mol·L-1KMnO4的毫升數表示H2O2酶活性;脲酶(Ure)采用苯酚-次氯酸鈉比色法,以培養1 d后1 g土壤中生成NH3-N 的毫克數表示脲酶活性;蔗糖酶(Suc)采用3,5-二硝基水楊酸比色法,以培養1 d后1 g土壤中生成葡萄糖的毫克數表示蔗糖酶活性;酸性磷酸酶(Acp)采用磷酸苯二鈉比色法,以1 g·h-1土壤中釋放的酚毫克數表示磷酸酶活性。每個土壤樣品至少測定3次,取平均值。
2.4 植物多樣性
植被多樣性指數采用如下測度[14],其計算公式如下:
Margalef豐富度指數(R):R= (S- 1)/lnN
Shannon-Wiener多樣性指數(H):
H= -∑(PilnPi)
Simpson多樣性指數(D):D= 1 - ∑(Pi/N)2
Pielou均勻度指(J):J=H/lnS
式中:Pi=Ni/N,即某個物種的相對多度;Ni為種i的株數;S為種i所在樣方的所有物種的總株數[15]。
2.5 數據處理
采用Microsoft Excel 2013對實驗數據預處理、數據統計和作圖;采用單因素方差分析(Oneway Anova)對各環境因子(水熱條件和植物多樣性、土壤理化性質)進行多重比較和差異性分析。采用前向選擇法對各環境因子進行約束性分析,然后使用蒙特卡洛檢驗對環境因子進行重要性排序;根據重要性排序的結果,篩選出對酶活性具有顯著影響的環境因子,采用Canoco5.0中的T-value雙序圖進行單一因子檢驗,明確其與各酶活性的關系,若某個土壤酶活性指標的箭頭連線落入或者穿過黑色圈表示呈顯著正相關,落入或穿過白色圈為顯著負相關[16]。
3 結果與分析
3.1 不同海拔植被帶水熱條件及物種多樣性特征
圖1表明:不同植被帶年均氣溫、年均降水量及土壤溫度沿海拔梯度的變化差異顯著(P<0.05),其中,年均氣溫和土壤溫度隨海拔升高而降低,即亞熱帶季風常綠闊葉林 > 半濕潤常綠闊葉林 > 濕性常綠闊葉林,年均降水量隨海拔升高而增加,即亞熱帶季風常綠闊葉林 < 半濕潤常綠闊葉林 < 濕性常綠闊葉林。
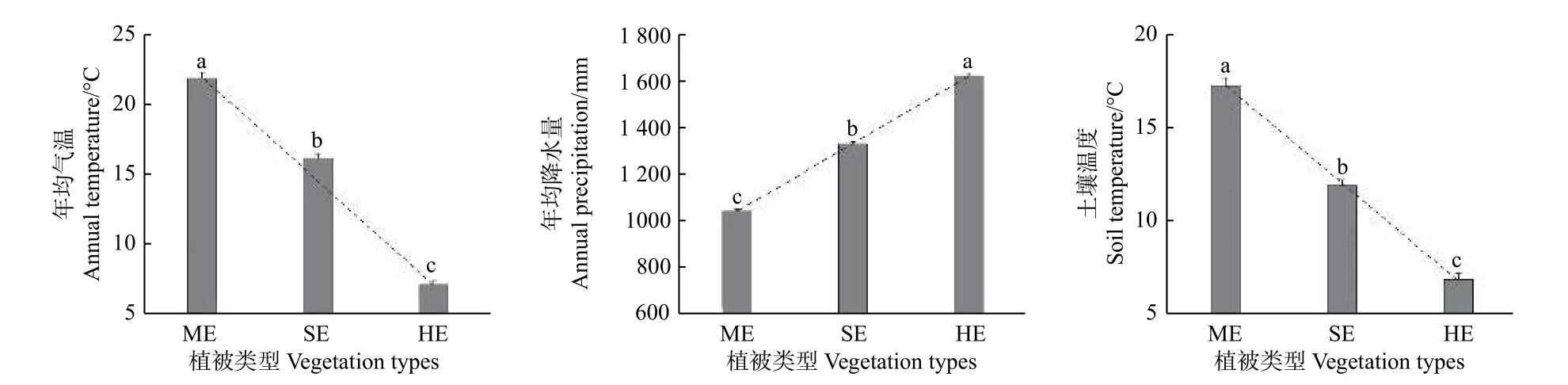
圖 1 不同植被帶水熱條件的海拔變化特征Fig. 1 The hydrothermal conditions of different vegetation zones at different altitudes
表2表明:不同植被帶Margalef(R)、Shannon-Wiener(H)、Simpson(D)和Pielou(J)指數沿海拔梯度的變化趨勢不同,R、H和J指數均隨海拔升高而減少,其中,3個植被帶的R、H指數差異顯著(P< 0.05),亞熱帶季風常綠闊葉林和半濕潤常綠闊葉林的J指數差異不顯著(P> 0.05);D指數沿海拔升高而增加,且3個植被帶間差異顯著(P< 0.05)。
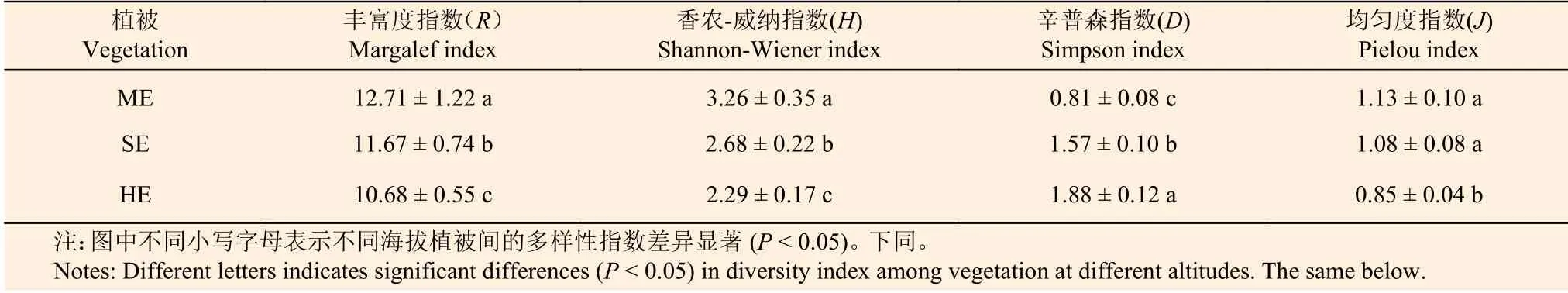
表 2 不同海拔樣地植物多樣性指數Table 2 The diversity index of plant communities at different altitudes
3.2 不同海拔植被帶土壤理化性質特征
3.2.1 土壤理化性質沿植被帶變化 由表3可知:各海拔植被帶土壤理化性質沿海拔梯度呈不同變化規律,且差異顯著(P< 0.05);土壤容重和pH值分別隨海拔升高而降低;土壤含水量、總有機碳、全氮、全鉀、速效氮和速效鉀隨海拔升高而增加,土壤全磷和速效磷隨海拔升高先降低后增加。濕性常綠闊葉林土壤含水量、總有機碳、全氮、全磷、全鉀、速效氮、速效磷和速效鉀的含量分別是亞熱帶季風常綠林的1.53、2.61、2.70、1.66、2.26、1.77、2.18、1.71倍。
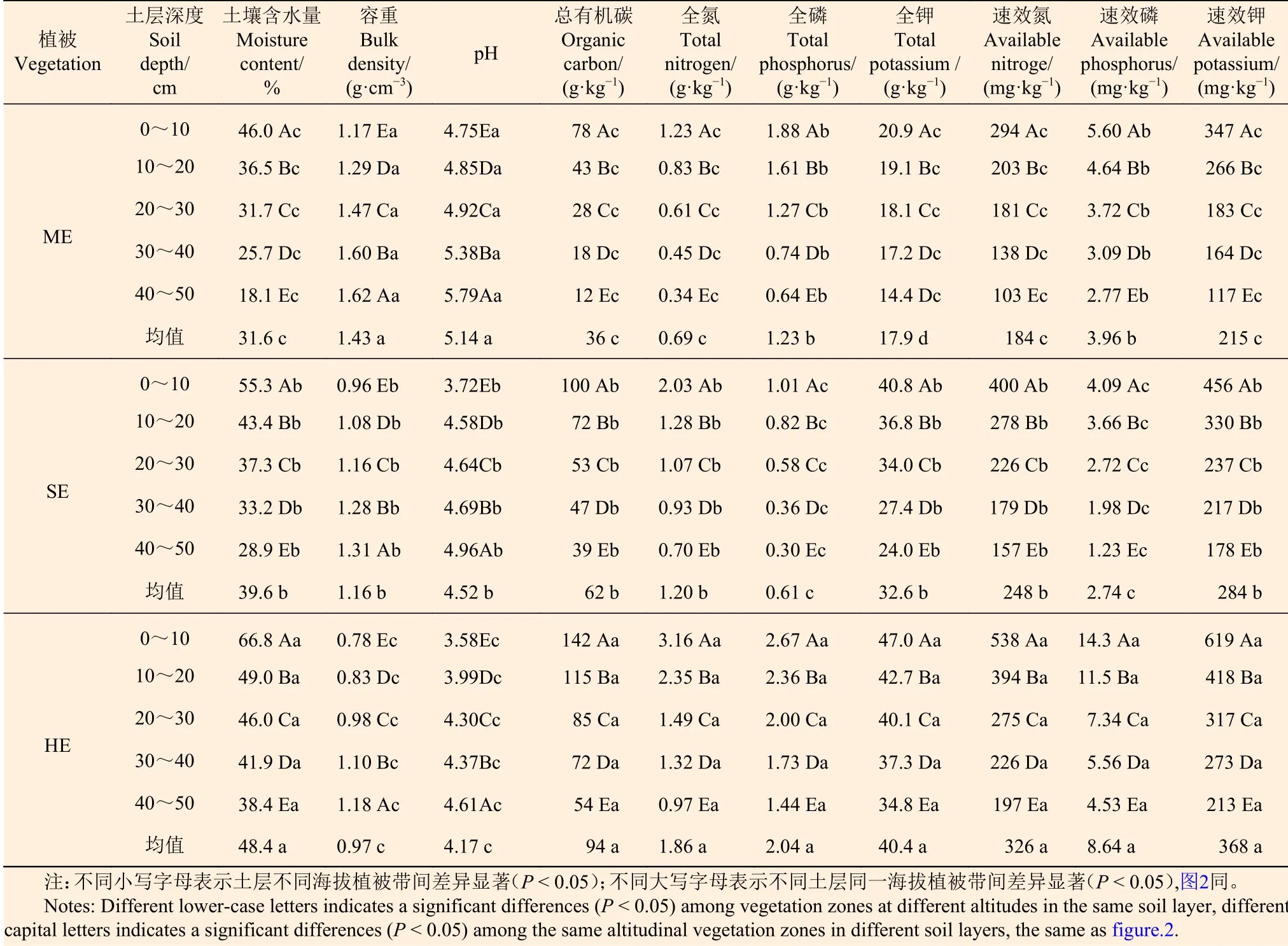
表 3 不同海拔植被帶土壤理化性質特征 Table 3 Soil physicochemical properties of vegetation zones at different altitudes
3.2.2 土壤理化性質的垂直變化 各植被帶土壤理化性質沿土層深度呈現不同變化規律,且差異顯著(P< 0.05)(表3)。土壤含水量、總有機碳、全氮、全磷、全鉀、速效氮、速效磷和速效鉀的含量均表現為隨土層加深而減小,0~10 cm最大,40~50 cm最小;與0~10 cm相比,40~50 cm的上述各指標,亞熱帶季風常綠闊葉林分別減小27.9 %、66 g·kg-1、0.89 g·kg-1、1.24 g·kg-1、6.5 g·kg-1、191 mg·kg-1、2.83 mg·kg-1、230 mg·kg-1,半濕潤常綠闊葉林分別減小26.4 %、61 g·kg-1、1.33 g·kg-1、0.71 g·kg-1、16.8 g·kg-1、243 mg·kg-1、2.86 mg·kg-1、278 mg·kg-1,濕性常綠闊葉林分別減小28.4 %、88 g·kg-1、2.19 g·kg-1、1.23 g·kg-1、12.2 g·kg-1、341 mg·kg-1、9.77 mg·kg-1、406 mg·kg-1。土壤容重和pH值均表現為隨土層加深而增加,40~50 cm最大,0~10 cm最小。
3.3 不同海拔植被帶土壤酶活性特征
3.3.1 不同植被帶土壤酶活性的比較 土壤脲酶、過氧化氫酶和蔗糖酶活性在不同海拔植被帶間差異顯著(P< 0.05),且隨海拔上升呈增大趨勢(圖2)。脲酶、過氧化氫酶和蔗糖酶活性均為濕性常綠闊葉林 > 半濕潤常綠闊葉林 > 亞熱帶季風常綠闊葉林。
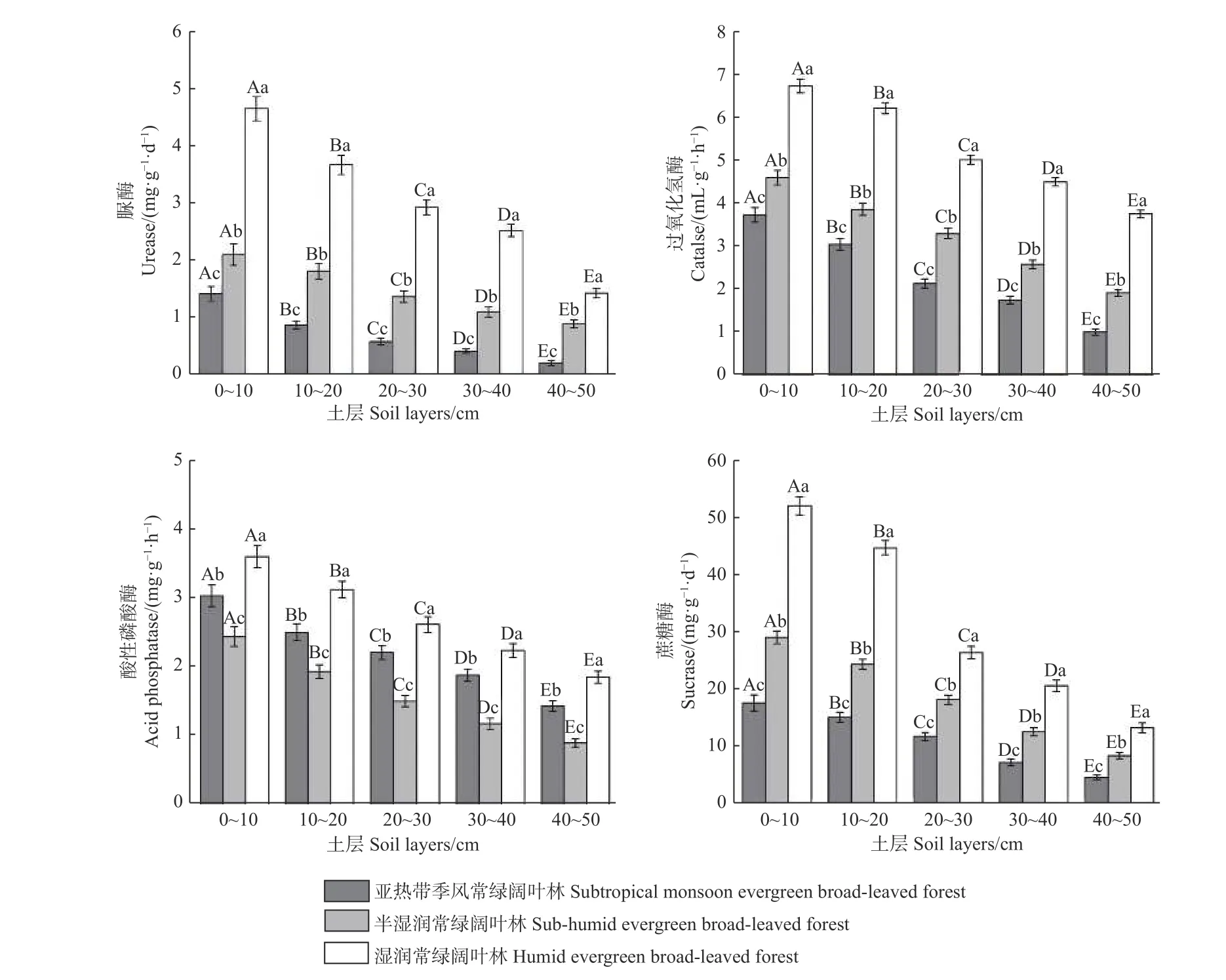
圖 2 不同海拔植被帶土壤酶活性變化Fig. 2 Soil enzyme activities of different vegetation zones at different altitudes
不同海拔植被帶酸性磷酸酶活性差異顯著(P<0.05),且隨海拔上升呈先減小后增大趨勢(圖2),其活性表現為濕性常綠闊葉林 > 亞熱帶季風常綠闊葉林 > 半濕潤常綠闊葉林。
3.3.2 土壤酶活性隨土層的變化 土壤酶活性沿剖面的變化均差異顯著(P< 0.05),且隨土層加深而降低,表現為0~10 cm土層最高,40~50 cm土層最低。土壤脲酶、過氧化氫酶、酸性磷酸酶、蔗糖酶其活性呈現:0~10>10~20>20~30>30~40>40~50 cm。同土層的酶活性沿海拔上升呈顯著不同的變化(P< 0.05),土壤過氧化氫酶、脲酶和蔗糖酶活性沿海拔升高呈增加趨勢,酸性磷酸酶活性呈先降低后增加趨勢。同一植被帶下,土壤酶活性均隨土層加深逐漸降低(圖2)。
3.4 土壤酶與環境因子的相關關系
3.4.1 環境因子對土壤酶活性的冗余(RDA)排序 水熱條件、植物多樣性、土壤理化性質等環境因子對4種土壤酶活性在第1排序軸解釋量為93.83%,第2排序軸解釋量為3.97%,即前2軸對酶活性特征解釋值為97.80%。表明前2軸能較好的解釋土壤酶活性與環境因子的關系,并且主要由第1排序軸決定。第1、第2排序軸中,水熱條件和植物多樣性因子與土壤酶活性的相關系數分別為0.999和0.998,進一步反映出土壤酶活性與環境因子關系密切。
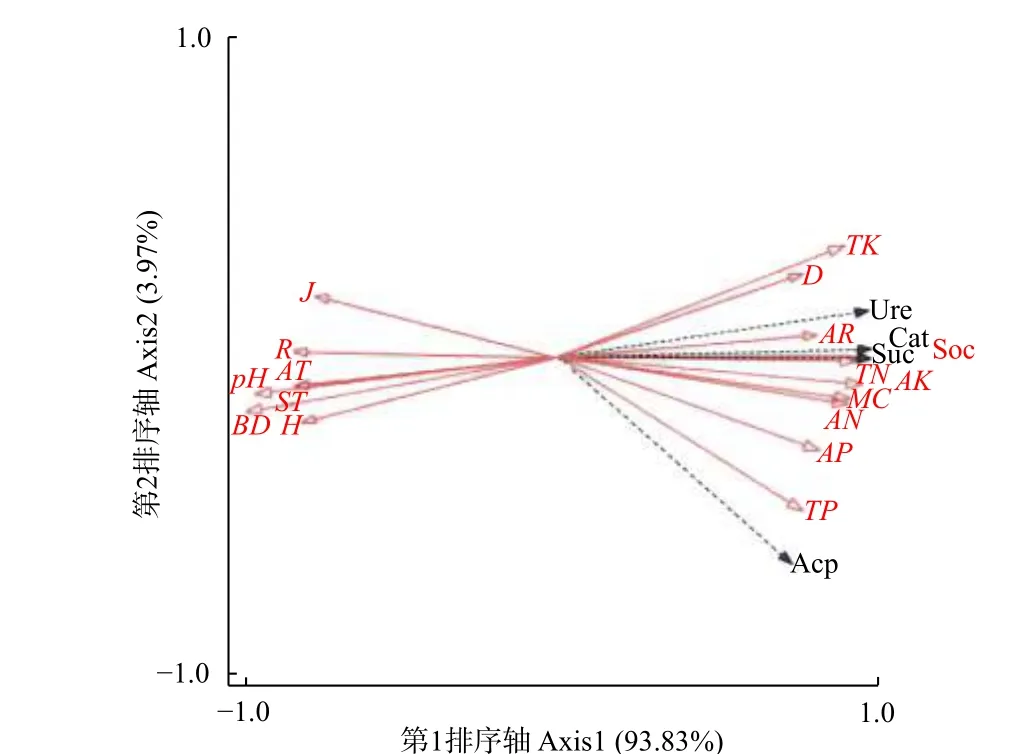
圖 3 土壤酶活性與環境因子的冗余度分析Fig. 3 Redundancy analysis of soil enzyme activity and environmental factors
從土壤酶活性與環境因子的二維排序圖(圖3)可知:土壤全鉀(TK)、pH和容重(BD)的箭頭連線較長,能較好解釋土壤酶活性的差異,其中,Margalef(R)、Shannon-Wiener(H)、Pielo-u指數(J)及年均氣溫(AT)、土壤溫度(ST)與過氧化氫酶(Cat)、脲酶(Ure)、蔗糖酶(Suc)和酸性磷酸酶(Acp)夾角較大(> 90°),呈顯著負相關(P< 0.05);Simpson指數(D)、土壤全鉀(TK)、速效鉀(AK)、含水量(MC)、速效氮(AN)、速效磷(AP)與上述4個酶夾角較小(< 90°),呈顯著正相關,其中,過氧化氫酶、蔗糖酶與總有機碳(SOC)和全氮(TN)夾角最小,脲酶與年均降水量(AR)夾角最小,酸性磷酸酶與全磷(TP)夾角最小,表明總有機碳和全氮對過氧化氫酶、蔗糖酶存在極顯著正效應(P<0.01),年均降水量對脲酶存在極顯著正效應,全磷對酸性磷酸酶存在極顯著正效應,而4種酶活性與pH和容重(BD)夾角最大,說明4種酶活性與pH和容重存在顯著負效應。
綜上可知:不同環境因子對土壤酶活性的影響差異較大。使用蒙特卡羅檢驗對17個因子進行大小排序得出:總有機碳(SOC)> 全氮(TN)> 容重(BD)> 速效鉀(AK)>pH> 含水量(MC)>速效氮(AN)> 全鉀(TK)> 有效磷(AP)> 全磷(TP)> Pielou指數(J)> 年均降水量(AR)>年均氣溫(AT)> 土壤溫度(ST)> Simpson指數(D)> Margalef index指數(R)> Shannon-Wiener指數(H)。相關分析顯示:各環境因子對土壤酶活性均影響顯著(P< 0.05)。
3.4.2 單一因子對酶活性的影響 根據蒙特卡洛檢驗排序的大小,使用T-value雙序圖篩選出對土壤酶活性有顯著影響的12個環境因子進行深入分析,以明確酶活性與主要環境因子的關系。由圖4a、b、d可知:脲酶(Ure)、過氧化氫酶(Cat)、蔗糖酶(Suc)和酸性磷酸酶(Acp)的箭頭均穿過了黑圈,說明土壤總有機碳(SOC)、全氮(TN)速效鉀(AK)與上述4個酶均呈顯著正相關,對各酶活性影響較大。由圖4f~h、l可知:過氧化氫酶、脲酶、蔗糖酶的箭頭穿過了黑圈,說明土壤含水量(MC)、速效氮(AN)、全鉀(TK)和年降水量(AR)和上述3個酶均呈顯著正相關,對3個酶活性變化起促進作用。由圖4c、e、k可知:脲酶、過氧化氫酶和蔗糖酶的箭頭均穿過了白圈,說明容重(BD)、pH和Pielo-u指數(J)與脲酶、過氧化氫酶和蔗糖酶呈顯著負相關,并對3個酶反向變化的影響較明顯。由圖4i、j可知:酸性磷酸酶箭頭完全落入了黑圈,說明速效磷(AP)和全磷(TP)與酸性磷酸酶呈極顯著正相關,表明速效磷和全磷是影響酸性磷酸酶的正向變化主要因子。由圖4c、l可知,酸性磷酸酶箭頭完全落入白圈,說明容重(BD)和年均降水量(AR)與酸性磷酸酶呈極顯著負相關,表明年均降水量和土壤容重對酸性磷酸酶反向變化的影響最明顯。
4 討論
4.1 不同植被帶土壤酶活性沿海拔的變化
高原山區垂直氣候沿海拔梯度的變化,能夠引起植被類型、凋落物輸入質與量、土壤微生物及理化性質的顯著改變,從而形成土壤酶的結構及活性在不同海拔高度的顯著差異性[17]。本研究中,土壤過氧化氫酶、脲酶及蔗糖酶的活性均隨海拔升高呈顯著增強的趨勢,且濕性常綠闊葉林的最大。濕性常綠闊葉林郁閉度最大,凋落物輸入量大,根系及分泌物較多、腐殖質層較厚,導致土壤碳、氮及磷等養分含量的顯著增加,其分解中釋放的土壤中有機質、養分元素可以為酶促反應提供良好的基質[18]。土壤過氧化氫酶與蔗糖酶隨海拔的增加,能夠促進土壤有機物的轉變與積累[19],土壤脲酶的增加能夠分解土壤中的尿素,提高土壤的供氮能力[20]。因此,土壤酶活性隨海拔的增加可能促進高海拔樣地土壤養分循環、土壤生物活動、植物生長及群落演替。
土壤酸性磷酸酶參與土壤磷循環和代謝,其活性的高低能夠反映土壤磷的生物轉換方向及強度[21]。本研究中,土壤酸性磷酸酶活性隨海拔升高呈先減小后增強的變化趨勢,這與任玉連等[9]研究結果不同,這可能與不同地區土壤空間異質性有關。半濕潤常綠闊葉林土壤磷含量顯著低于亞熱帶季風常綠闊葉林和濕性常綠闊葉林,表明受到的磷限制程度低于其它2種植被帶,無需微生物分泌更多的磷酸酶來維持速效磷供給[22]。因此,土壤酸性磷酸酶活性沿海拔的變化反映了植物生長需求及土壤供磷水平之間的平衡關系。
不同植被帶相同土層酶活性隨海拔上升呈增加趨勢。一方面,隨著海拔上升,同一土層容重變小,根系分布逐漸增多,植物根系分泌物的增加促進土壤微生物的代謝產酶能力[23];另一方面,相同土層酶活性的海拔變化規律與養分元素一致,表明高海拔植被帶土壤剖面的水熱條件和肥力狀況良好,促進了土壤酶活性。在土壤層次上,由于受生物、淋溶等共同作用,使土壤中養分物質在不斷地進行交換,導致各土層間土壤理化性質的顯著差異[24],從而對土壤酶活性的垂直分布造成影響。研究發現,各海拔植被帶下4種土壤酶活性沿土層逐漸減小,這與土壤含水量和土壤碳、氮和磷等養分含量垂直變化趨勢相一致。因此,不同海拔土壤酶活性垂直變化與土壤微生物及土壤養分沿土層分布密切相關。
4.2 不同海拔水熱條件及植物多樣性對土壤酶活性的影響
海拔是重要的山地地形因子之一,海拔的變化會引起水熱條件的梯度變化[25],Zuccarini等[26]研究發現,土壤水分會對酶活性產生顯著的影響,升溫時只有伴隨充足的水分條件才能增加土壤酶活性。本研究發現,降水量和土壤含水量對脲酶、過氧化氫酶和蔗糖酶存在顯著正向影響,但降水量的增多會對酸性磷酸酶產生顯著負效應。這與酶的底物變化有關,水分沿海拔的增加能夠促進凋落物分解和高分子化合物形成[27],積累的有機質增加了土壤脲酶、過氧化氫酶和蔗糖酶的反應底物,但大量降雨加劇了土壤磷素的淋溶作用,導致研究區整體處于低磷狀態(全磷均值為1.29 g·kg-1),使磷酸酶對磷的底物利用率較低。前人研究表明[28-29],溫度可直接或間接影響酶活性,適宜的溫度對酶活性具有促進作用。本研究“環境因子對酶活性影響的排序”表明,土壤溫度對酶活性總體解釋性較弱,低海拔植被帶較高的土壤溫度未能顯著影響到土壤酶的活性。可能由于文山低海拔植被帶日照時間長,強光合作用促進植被呼吸速率[30],導致生境熱量循環加快,刺激了微生物對土壤碳氮等酶促底物的分解速率,進而限制低海拔植被帶土壤酶活性。
植被多樣性不僅影響著森林群落結構組成,還對土壤基質及養分庫起穩定作用[31],從而引起土壤酶活性對植被多樣性的變化做出響應。本研究中,植被多樣性沿海拔梯度對土壤酶活性產生不同程度的負向影響,其中,均勻度指數最明顯,這與楊秉珣等[32]研究結果相似。保護區主要以典型亞熱帶闊葉林為主,群落組成差異較小,均勻度指數對群落分布的影響較其他指數強,隨著海拔上升,植被由草本、灌木群落向喬木群落進行演替,地表覆蓋物和地下根系腐解物逐漸增多,伴隨著低溫環境,微生物的呼吸作用和分解能力受到抑制,土壤養分物質以積累為主[33],從而使土壤酶的底物和活性隨海拔上升均有不同程度的增加。
4.3 不同海拔植被帶土壤理化性質對土壤酶活性的影響
土壤理化性質影響著土壤酶的活性及穩定性[34],但不同土壤理化因子對土壤酶活性影響程度有差別。土壤有機質和礦質元素是土壤中酶促底物的主要供源,是土壤酶的載體[35]。本研究中,土壤有機質和礦質元素(氮、磷、鉀)對土壤各酶活性存在顯著的正效應,其中,總有機碳和全氮最明顯,表明土壤養分沿海拔的增加促進了樣地酶活性,這與許全等[36]研究結果相似。
土壤pH能夠決定土壤有機分子功能基團的離子化,影響酶和底物的構象[28],本研究發現,土壤pH與各個酶活性呈顯著負相關,表明土壤酶活性隨土壤pH的升高而降低,這與Sinsabaugh等[37]等研究結果不同,這可能是由土壤的空間異質性,造成土壤pH和酶活性的關系不同所致。土壤容重通過改變土壤緊實度來影響土壤水分條件、空氣組成和熱量狀況,進而調控土壤酶活性[38]。研究表明,土壤容重與各酶活性呈顯著負相關,表明土壤酶活性隨土壤容重增加而降低,這與呂瑞恒等[6]和馬劍等[7]研究結果相似。
土壤理化性質的海拔差異對土壤酶活性分布規律產生重要影響,土壤酶活性的增強,則能夠改變土壤理化性質,有利于土壤養分元素的積累,促進植物生長及群落演替[39]。因此,文山國家級自然保護區“海拔高度-植被群落-土壤理化性質-土壤酶”之間存在復雜的耦聯關系。
5 結論
不同植被帶土壤酶活性沿海拔梯度呈現不同的變化特征,較高海拔的濕性常綠闊葉林土壤酶活性顯著高于較低海拔的亞熱帶季風常綠闊葉林與半濕潤常綠闊葉林群落。土壤過氧化氫酶、脲酶和蔗糖酶活性隨海拔升高呈增加趨勢,而土壤酸性磷酸酶活性則呈先下降后上升的趨勢。不同海拔植被帶各土壤酶活性均隨土層加深而降低。
冗余分析和單一因子檢驗表明,各環境因子對酶活性具有不同的影響。土壤有機質和礦質元素(氮、磷、鉀)是調控土壤酶活性變化的主要因子;容重和pH與土壤酶活性呈顯著負相關,表明較低的容重和pH可能有利于提高土壤酶的活性;植被多樣性和水熱條件的海拔差異是驅動酶活性變化的重要因子。
