驅動蛋白分子KIF18A的結構與功能研究進展
2020-12-14 06:14:04朱長軍周之春
天津師范大學學報(自然科學版) 2020年6期
朱長軍 ,周之春
(1.天津師范大學生命科學學院,天津 300387;2.天津師范大學分子細胞系統生物學重點實驗室,天津300387;3.天津師范大學天津市動植物抗性重點實驗室,天津 300387)
細胞分裂是生命個體存活、生長以及物種繁衍的基礎,是母細胞分裂成子細胞的過程.有絲分裂是最主要的真核細胞分裂方式.有絲分裂過程中,紡錘體微管的精準調控、微管與動粒的附著(KT-MT attachment)是染色體整列(chromosome alignment)和中板集合的核心[1-2].細胞在有絲分裂前中期(prometaphase),高度動態不穩定的動粒微管能夠有效捕獲染色體[3],牽引著染色體向赤道板聚集整列.有絲分裂中期,動粒微管協助染色體整齊地排列在紡錘體中間區域(亦稱染色體中板集合),紡錘體組裝檢驗點(spindle assembly checkpoint,SAC)嚴格監控染色體的中板集合過程.有絲分裂后期,SAC失活,姐妹染色單體彼此分離并移向紡錘體兩極,最終進入2個子細胞中.
驅動蛋白分子是以微管為軌道的馬達蛋白,在細胞器遷移、組織器官發育、信號傳導、有絲分裂、減數分裂等過程中發揮了重要作用.驅動蛋白kinesin-8家族的多種微管結合蛋白(microtubule-associated protein,MAP)通過影響微管的聚合和解聚行使著調控微管動態不穩定性的功能.KIF18A是kinesin-8家族的一員,能夠以微管為軌道向正極運動,傾向于結合較長的微管,其活性具有長度依賴性并影響著紡錘體的長度,可保障姐妹染色體整列的及時順利完成[4-6].本文就KIF18A的蛋白結構、細胞內定位及其在有絲分裂過程中的調控作用等加以綜述.
1 KIF18A基因和蛋白分子的結構與功能
1.1 KIF18A基因及其蛋白結構
2005年,Luboshits等[7]在人的骨髓間充質干細胞表達文庫中克隆出了MS-KIF18A,它在人體的睪丸和氣管中均有表達.該基因屬于kinesin-8家族,定位于人類染色體11p14,包含17個外顯子,基因全長約87.57kb,開放閱讀框(open reading frame,ORF)2 697 bp,編碼898個氨基酸,蛋白相對分子質量約為102×103.MSKIF18A的蛋白分子功能域如圖1所示.

圖1 KIF18A蛋白分子的結構和運動Fig.1 Structure and movement of KIF18A protein
MS-KIF18A的功能域包括:N端球形的馬達結構域(motor domain),位于氨基酸序列N9到N363,其中ATP結合催化結構域為N333~N340,微管結合位點結構域為N304~N308,賦予其靶向微管正極的運動能力及類似微管解聚酶的活性;中央卷曲螺旋結構域(helical coiled-coil domain),位于氨基酸序列N375到N463,主要由α-螺旋形成同源二聚體;C端扇形的尾部結構域(tail domain),位于氨基酸序列N464到N898,具有微管結合位點,可調控KIF18A蛋白功能,使其能夠穩定運動到微管正極末端并聚集[8-9],同時也含有多個蛋白激酶作用的保守序列.在N端和桿狀區之間有一段Neck鉸鏈區域,位于氨基酸序列N364到N374,與驅動蛋白13家族的微管解聚酶結構域序列一致,Neck鉸鏈的長度保障了KIF18A在微管上的運動和穿過微管結合蛋白障礙物的能力[10].KIF18A蛋白以二聚體形式在微管上向微管正極末端運動,細胞周期依賴性激酶1(CDK1)可對其674和684位點的絲氨酸進行磷酸化修飾.
KIF18A蛋白在多個物種中均有表達.應用TCOFFEE和MEGA7.0軟件對不同種屬間的KIF18A蛋白進行序列比對和進化樹分析,結果分別如圖2和圖3所示.由圖2和圖3可以看出,Kif18A蛋白家族的分子結構在不同物種中非常保守,特別是該蛋白分子的馬達結構域.這些現象預示該蛋白分子在生命進化過程中調控細胞的基本生命活動:一方面,該蛋白分子的功能與生命進化過程中細胞的基本功能密切相關;另一方面,該蛋白分子在不同物種中的功能應該非常相似,具有保守性.因此,人們可以使用多種模式生物,如酵母、果蠅、非州瓜蟾等研究KIF18A蛋白分子的功能及其應用,還可以應用小鼠等動物模型為以Kif18A蛋白為靶點的藥物開發提供前期實驗指導.
1.2 KIF18A基因的轉錄
KIF18A與雌激素受體ERɑ在體外和體內均可結合,雌激素水平較高的細胞中KIF18A在mRNA水平和蛋白水平上都有所增加.KIF18A基因的轉錄可通過轉錄因子雌激素受體ERα被雌激素激活,受雌激素正調節[11-13].應用染色質免疫沉淀實驗和熒光素酶報告系統檢測發現,這些細胞中轉錄因子ERα和c-Jun均可以結合并活化KIF18A基因的啟動子,如圖4所示,ERα是負責KIF18A基因轉錄的主要轉錄因子[11].KIF18A的啟動子區有多個轉錄因子結合位點,在ERɑ缺失細胞中是否存在其他轉錄因子與KIF18A啟動子結合調控KIF18A基因的轉錄,還有待深入研究.
1.3 KIF18A蛋白分子的細胞內定位
KIF18A是細胞周期特定時期的表達蛋白[4]:G1期細胞內KIF18A的表達極低,G2/M期KIF18A表達明顯提高.在間期細胞(interphase)中,由于C端存在核定位序列,因此KIF18A主要位于細胞核內且與微管正極結合蛋白(End-Binding 1,EB1)共定位,即定位于微管正極末端.在有絲分裂前中期和中期細胞中,KIF18A主要定位在微管正極末端和動粒外板間,也可在紡錘體微管和星體微管檢測到KIF18A染色.后期和末期細胞中,KIF18A蛋白轉移至紡錘體中央區(spindle midzone)和中小體(midbody),少量定位在紡錘體和星體微管正極末端[14].KIF18A蛋白分子的馬達區和尾區均有微管結合的功能區,尾區含核定位序列.馬達區和桿狀區共同作用才能使該蛋白在有絲分裂末期定位于中體.KIF18A蛋白分子在細胞內的定位還可能受蛋白翻譯后修飾如磷酸化修飾等的調控.

圖2 不同物種間KIF18A蛋白分子序列的比對Fig.2 Sequence alignment of KIF18A proteins among different species

圖3 KIF18A蛋白家族在不同物種中的進化分析Fig.3 Evolutionary analysis of KIF18A protein family in different species
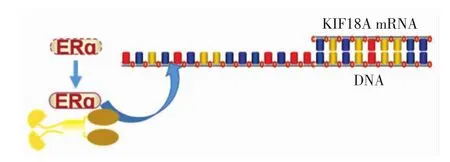
圖4 KIF18A和雌激素受體ERα相互結合激活KIF18A基因的轉錄Fig.4 Interaction between KIF18A and ERα(estrogen receptor)and activativation on the transcription of KIF18A gene
1.4 KIF18A蛋白分子的功能
1.4.1 KIF18A的運動功能(motor activity)
KIF18A的馬達結構域位于N端,在C端存在微管結合位點.KIF18A的運動以微管為軌道,體外微管滑動實驗(microtubule-gliding assay)結果發現,全長KIF18A可向微管正極運動[4,15],這表明KIF18A與其在果蠅和酵母中的直系同源物(Klp67A和Kip3)一樣,均為靶向微管正極運動的馬達蛋白[6].細胞中的KIF18A抗體免疫熒光染色結果表明:在有絲分裂早期,內源性KIF18A與動粒外板蛋白Hec1共定位;中期,外源GFPKIF18A與動粒(CREST)共定位.GFP-KIF18A-MD突變體(MD,motor dead,馬達結構域突變失去馬達活性)的免疫熒光染色結果顯示,GFP-KIF18A-MD沿紡錘體微管彌散分布,不與動粒共定位,這說明KIF18A依賴其N端的馬達結構域在微管上向正極滑動并最終定位到動粒微管末端[5].
1.4.2 KIF18A的微管解聚酶活性(depolymerase activity)
KIF18A在酵母中的直系同系物kinesin-8家族成員kip3能夠以長度依賴方式發揮微管解聚酶的活性[6].在人的HeLa細胞中,應用小分子RNA干擾KIF18A的表達后細胞內微管的穩定性明顯升高,抑制了冷誘導的微管解聚[4].純化的KIF18A蛋白在體外實驗中能夠以ATP依賴的方式直接解聚微管,且對長微管的解聚能力明顯高于短微管[4].關于KIF18A的微管解聚酶活性尚有爭議.Du等[15]報道KIF18A是通過抑制微管蛋白在正極的聚合從而抑制微管伸長,并非直接解聚微管.在微管動態不穩定性中,KIF18A并不影響災變(catastrophe)的頻率,但會使修復(rescue)的頻率增加,進而使得整體的停滯時間延長.因此KIF18A可能通過兩種方式抑制微管正極的動態不穩定性:解聚微管或阻止新的微管蛋白組裝[16].
2 KIF18A的生物學作用及其調控機制
2.1 KIF18A在有絲分裂中的作用
KIF18A利用N端含有的ATP結合位點,依靠水解ATP供能在微管上運動并運送與其C端結合的貨物.通過與定位在生長微管正極末端的EB1蛋白進行共定位研究,確定KIF18A是一個正極朝向的驅動蛋白[17].KIF18A在細胞周期中的表達具有特異性,在G1期表達較低,在G2/M期表達量升高[4],這表明KIF18A主要在細胞有絲分裂期發揮功能.如圖5所示,KIF18A可促進染色體排列到赤道板上,為后續姐妹染色單體的精確分離做準備,保證細胞有絲分裂的正常進行[4-5,17-20].
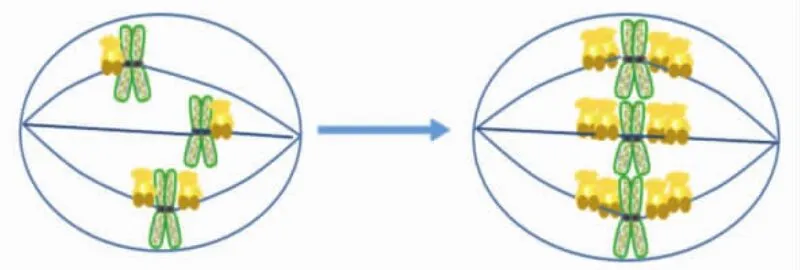
圖5 KIF18A在有絲分裂早期促進姐妹染色體的中板集合Fig.5 KIF18A promotes congression of sister chromosomes in early mitosis
抑制細胞內KIF18A的表達會導致姐妹染色單體移動速率加快,動粒振幅(oscillation amplitude)增大,染色體轉向頻率(switch rate)降低,同時動粒內部間距(inter-kinetochore distance,IKD)增大,張力降低,紡錘體變形(拉長形成香蕉形紡錘體)[4].細胞轉染過表達GFP-KIF18A融合蛋白后的染色結果顯示紡錘體縮短,動粒內部間距變小,染色體振幅降低,染色體震動頻率降低[6].這些現象說明KIF18A可降低染色體的振動頻率,縮短動粒振幅,促進染色體轉向,抑制紡錘體延伸,降低紡錘體的動態不穩定性,而這些作用都依賴于KIF18A蛋白具有的馬達活性.
有絲分裂后期細胞中單個核的形成也需要KIF18A發揮正常功能.Fonseca等[21]發現,依賴KIF18A的中期染色體排列使染色體到達極的距離相等,KIF18A缺失細胞中的落后染色體在后期移動距離則較長.KIF18A缺失會導致間期細胞核組織缺陷,后期核形態異常,末期核纖層蛋白分布失常,p53依賴性檢驗點則能夠抑制形成微核的KIF18A缺陷細胞進行分裂[21].
利用KIF18A的小分子化合物抑制劑BTB-1處理細胞時,也可觀察到“香蕉狀”紡錘體,因此認為KIF18A對于紡錘體形態的維持具有重要作用[22].抑制KIF18A的表達或功能可將細胞阻滯于有絲分裂早期,并激活細胞有絲分裂紡錘體檢驗點.在缺失KIF18A的細胞中,對Mad2的染色定位表明,染色體未正確連接微管時SAC檢驗點會被激活.Zhu等[17]、Janssen等[23]研究發現,細胞缺失KIF18A會導致微管與動粒正常連接處動粒之間的張力異常,從而激活有絲分裂紡錘體檢驗點,進而影響細胞的有絲分裂進程.
2.2 KIF18A對精子發育的作用
KIF18A對于細胞分裂過程中正常的染色體分離是必不可少的.Liu等[24]研究發現,KIF18A功能缺失會引起微管動力學、紡錘體完整性和檢驗點激活的嚴重缺陷,從而導致小鼠發生性細胞再生障礙.敲除KIF18A基因表達可導致雄性小鼠完全不育,但雌性小鼠沒有受到影響.組織學檢驗結果顯示,KIF18A-/-小鼠的睪丸生精小管發育嚴重受損,從而導致小鼠睪丸萎縮,在有絲分裂和減數分裂過程中造成染色體重組缺陷.進一步研究表明,KIF18A基因沉默會導致CENP-E和BubR1降解并伴隨著姐妹染色單體的過早分離.這說明KIF18A與BubR1和CENP-E能夠在生理上相互作用并在有絲分裂過程中被調節.
Czechanski等[20]研究發現,生殖細胞缺陷 2(gcd2)突變小鼠在胚胎發育過程中出現生殖細胞缺失從而導致不育,這是KIF18A的馬達結構域中R308K的錯意突變造成的結果.KIF18A缺失會導致染色體排列缺陷,體細胞還可進行有絲分裂,而生殖細胞則會發生有絲分裂停滯和凋亡,這表明KIF18A是生殖細胞中細胞周期進展所必需的基因之一.
2.3 KIF18A生物學作用的調控機制
2.3.1 蛋白結構功能域對KIF18A功能的調控
KIF18A在有絲分裂間期主要定位于細胞核內,有絲分裂期沿紡錘體微管運動到動粒微管正極末端與動粒外板之間.Stumpff等[25]對野生型KIF18A進行截短突變實驗,發現僅N端結構域截短的突變體沿紡錘體微管分布,不能與動粒共定位;僅C端結構域截短的突變體可穩定結合微管,并能定位在細胞核內.兩種截短突變體都失去了調控紡錘體長度和染色體中板集合的功能,說明KIF18A的N端和C端均具有微管結合區,并且KIF18A在動粒的準確定位是其功能發揮的必要條件[8-9,25].KIF18A的N端截短突變(aa1-406)在體外的運動速率是全長野生型KIF18A的2.5倍,C端尾部截短則可減緩KIF18A的移動速率.進一步研究發現,C端尾部可通過靜電作用與微管結合,抑制KIF18A從微管解離,但KIF18A向微管正極運動以及對于微管動態不穩定性的調控仍然依賴N端[9,25].此外,Kim等[26]應用KIF18A尾部結構域與其他驅動蛋白(KIF4A、KIF5B)的馬達結構域構建融合蛋白發現,KIF18A在微管正極末端的定位及其對微管長度的調控不僅依賴尾部結構域,其馬達區的Loop2結構也發揮了關鍵作用.總之,KIF18A的功能發揮需要它的正確定位和分子馬達的活性,而其恰當定位又依賴于C端尾部與微管的結合以及N端馬達結構域的運動功能.2.3.2 蛋白翻譯后修飾對KIF18A功能的調控
KIF18A功能的發揮也受到翻譯后修飾的調控.細胞周期依賴激酶 1(cyclin-dependent kinase-1,CDK1)能夠催化KIF18A的Ser674與Ser684位點發生磷酸化.CDK1對KIF18A進行磷酸化后,動粒的振幅增加,這與敲除KIF18A的結果相似.細胞中轉入外源模擬磷酸化KIF18A后,磷酸化狀態的KIF18A無法運動到動粒微管末端并聚集,而是定位在微管網架上,因此磷酸化作用很可能會使KIF18A無法定位到微管末端,從而無法發揮降低微管動態不穩定性的功能,使得動粒振幅增大.H?fner等[27]通過活細胞成像實驗發現,KIF18A的CDK1磷酸化修飾并不會影響姐妹染色單體的中板集合(congression)時間,但延遲了中板變薄(thin metaphase plate,tMP)的時間,推測是 CDK1的磷酸化修飾影響了KIF18A的運動能力,進而影響了形成tMP的時間.De Wever等[28]發現蛋白磷酸化酶(PP1,protein phosphatase-1)可以特異性地結合并去磷酸化KIF18A,與蛋白激酶CDK1的作用相互拮抗,二者可共同調控KIF18A促進染色體整列和中板集合的活性.除CDK1外,KIF18A還與許多有絲分裂蛋白激酶具有共定位,如Aurora A、Aurora B、PLK1、Mad2 和MSP10等,這些蛋白激酶是否也參與磷酸化修飾并調節KIF18A的定位和功能尚不清楚.
KIF18A是細胞G2/M期特異性表達的蛋白分子,這預示著它在有絲分裂后期極有可能發生蛋白分子降解.Yang等[29]研究發現,KIF18A可被SUMO2共價修飾,雖然該修飾不影響KIF18A的蛋白穩定性和胞內定位,但非SUMO2化突變體會引起細胞周期阻滯,使得細胞無法離開有絲分裂期.KIF18A還是APC/C(the anaphase promotingcomplex/cyclosome)-Cdc20 的底物,如圖6所示,APC/C可以結合KIF18A的C端LR結構域(在453~653之間),在有絲分裂后期促使KIF18A降解[30].這些結果顯示,KIF18A的蛋白降解對于細胞有絲分裂與胞質分裂的順利進行和完成至關重要.
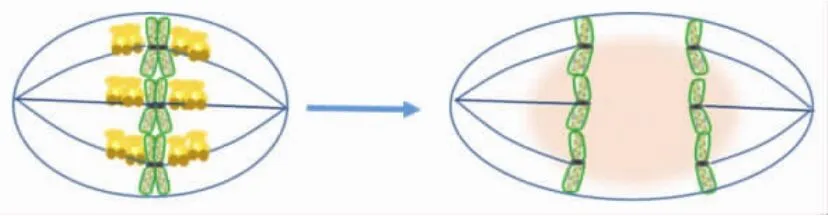
圖6 KIF18A在有絲分裂后期被降解Fig.6 Degradation of KIF18A at anaphase in late mitosis
2.3.3 生物大分子對KIF18A功能的調控
Ye等[31]研究發現,KIF18A可與多種蛋白分子相互作用,這對于KIF18A及其結合蛋白功能的正常發揮非常重要.如肝癌上調蛋白(hepatoma upregulated protein,HURP)可與KIF18A直接結合,促進KIF18A蛋白翻轉(turnover),抑制KIF18A向微管正極運動的能力,從而增加微管的穩定性.Huang等[32]的免疫共沉淀實驗結果表明,有絲分裂期的細胞中,KIF18A能夠與微管正極結合馬達蛋白CENP-E和有絲分裂檢驗點蛋白BubR1結合,同時KIF18A能夠提高CENP-E蛋白的穩定性,促進CENP-E在動粒處的定位,穩定動粒微管與染色體的結合.如圖7所示,微管結合蛋白KBP也可以結合KIF18A并抑制其活性.KBP能夠阻止KIF18A和KIF15的催化結構域與微管晶格相互作用,從而阻斷它們的活性.中期細胞中KBP過表達會抑制染色體的正常排列,降低紡錘體的長度,破壞紡錘體的雙極性,這些作用與KIF18A和KIF15的功能抑制是一致的.Malaby等[33]通過研究KBP過表達對KIF18A和KIF15定位的影響,也證實了KBP會干擾馬達蛋白之間的相互作用.

圖7 微管結合蛋白KBP與KIF18A相互作用并抑制KIF18A功能Fig.7 Microtubule binding protein KBP binds KIF18A and inhibits its function in early mitosis
Manning等[34]使用siRNA干涉細胞內源KIF18A后抑制了Astrin(aster-associated protein)在動粒處的定位,說明KIF18A可能在一定程度上通過促進Astrin在動粒的定位穩定KT-MT附著,進而調控染色體的中板集合[34].KIF18A還與Kid(kinesin-10家族蛋白)共同促進了染色體在赤道面的定位,而KIF4(kinesin-4家族蛋白)則與Kid一起協助染色體的整列[35].
HEST(kinesin-14家族蛋白)是一種調控紡錘體微管交聯與滑動的驅動蛋白.Weaver等[8]研究發現,干涉HEST會造成紡錘體縮短,同時干涉HEST與KIF18A則可以恢復由于KIF18A沉默引起的紡錘體異常延伸,這說明具有類似于解聚酶活性的KIF18A和具有滑動活性的微管結合蛋白HEST協作,可共同調控紡錘體微管的長度.
另外,KIF18A還參與調節Sirt2介導的微管蛋白的乙酰化.Tang等[36]研究發現,敲除KIF18A后卵母細胞減數分裂過程中乙酰化酶Sirt2的表達降低,乙酰化微管蛋白水平升高,最終導致紡錘體組織缺陷.微管蛋白K40R mRNA可誘導微管蛋白去乙酰化,保護卵母細胞免受KIF18A下調的影響,使得卵母細胞的紡錘體形態恢復正常.
3 KIF18A與腫瘤發生、發展的關系
KIF18A蛋白在多種腫瘤中均有高表達,包括肝細胞癌[37]、膠質母細胞瘤(Glioblastoma,GBM)[38]、乳腺癌[17]、肺癌[19]、膽管癌(CCA,cholangiocarcinoma)[39]、卵巢癌[40]、滑膜肉瘤(SynSa,Synovial sarcoma)[41]、橫紋肌肉瘤(RMS,Rha-bdomyosarcoma)[42]、前列腺癌[43]等,這預示著KIF18A與腫瘤的發生、發展密切相關,可成為多種腫瘤分子診斷和治療的靶標.
KIF18A的表達與臨床大腸癌發展有關.結腸癌是一種常見的實體癌,近年來全球發病率逐年增加[44-45].目前結腸癌治療的難點是預后,25%的結腸癌初發病人都有復發[46],復發后25%的病人出現轉移[43,47].Nagahara等[48]對113例初期CRS手術病人進行調查研究,發現這些病人中的KIF18A均明顯高表達,并與腫瘤分期、淋巴和血管遷移、淋巴轉移、組織浸潤相關,說明KIF18A與結腸癌的進展有關.Zhu等[49]、Zhong等[50]研究發現,KIF18A能夠誘導Akt磷酸化,小鼠中敲除KIF18A可明顯促進細胞凋亡,推測KIF18A能通過激活PI3K-Akt信號通路促進結直腸癌的發生發展.
KIF18A在人乳腺癌細胞中也高表達,且其過表達與乳腺腫瘤的分級、遷移和預后有關.Zhu等[17]、Alfarsi等[51]對乳腺癌細胞進行研究,發現過表達KIF18A會導致多核細胞的產生,低表達在體內外則均能大幅度減弱細胞增生能力,這是通過KIF18A在微管末端穩定微管以及PI3K-Akt信號轉導途徑的失活而誘導了細胞凋亡.Zhong等[50]、Li[52]等研究發現,肺腺癌中KIF18A在轉錄和翻譯水平均上調,KIF18A的異常表達與臨床病理惡性程度相關.在肺腺癌中能夠觀察到KIF18基因突變,其表達也受DNA拷貝數的調控,KIF18A基因敲除能夠抑制肺腺癌細胞的體內和體外增殖,誘導細胞凋亡和G2/M期阻滯.與KIF18A同時高表達的基因都集中在細胞周期信號通路中,因此深入研究KIF18A在肺腺癌中的作用機制具有重要臨床意義.
4 結論與展望
KIF18A是一個以微管為軌道向微管正極末端運動的分子馬達蛋白,通過影響微管末端動態不穩定性調控染色體的中板集合,在有絲分裂期發揮功能.在有絲分裂后期,該蛋白被泛素化降解,保證有絲分裂過程中染色體的精確分離,促使有絲分裂及胞質分裂的順利完成.有絲分裂早期,KIF18A在靠近動粒的微管正極末端定位是其功能發揮的必要條件,定位不僅依賴其N端的馬達活性,具有微管結合能力的尾部結構域也起到關鍵作用.KIF18A還受到可逆的蛋白磷酸化/去磷酸化修飾,但該蛋白翻譯后修飾如何調控KIF18A的功能尚缺乏系統研究.ERɑ可與KIF18A結合并促進其轉錄,但KIF18A是否還受其他轉錄因子的調控仍不清楚,因此KIF18A的基因轉錄調控機制研究尚待深入.在減數分裂過程中,缺少KIF18A的細胞將不能完成減數分裂,進而會導致雄性動物精子形成障礙和睪丸發育不良.在針對多種腫瘤的研究中都發現KIF18A與腫瘤的遷移、發生、發展、預后等有關,但其影響腫瘤遷移的機制尚不明確.鑒于KIF18A在腫瘤發生發展過程中的重要作用,以KIF18A為分子靶點探討多種腫瘤的早期診斷和有效治療將為不同腫瘤的診療提供新的思路和策略.
