大南湖二礦3種喬木的光合特性
2020-12-23 05:41:45王夢李云馬伊琍羅艷麗張繼衛時威麻浩
綠色科技 2020年14期
王夢 李云 馬伊琍 羅艷麗 張繼衛 時威 麻浩

摘要:為探究人工種植植被在當地的適應能力,為礦區生態修復提供理論依據。利用CIRAS-2型便攜式光合儀測定了新疆哈密大南湖二礦榆樹(Ulmus pumila L.)、沙棗(Elaeagnus angustifolia L.)、竹柳(Salix fragilis)這3種人工喬木的凈光合速率、蒸騰速率日變化,以及對影響植物凈光合速率的環境因子進行相關性分析,結果表明:①在野外自然條件下,7月的3種植物凈光合速率日均值大小表現為:竹柳>沙棗>榆樹,竹柳表現出有較強的光合作用;②影響榆樹、竹柳、沙棗出現光合“午休”現象的原因分別為非氣孔因素、共同作用、氣孔因素;③3種植物日均蒸騰速率由大到小依次為沙棗>榆樹>竹柳;④蒸騰速率與胞間CO2濃度是影響植物凈光合速率的主要環境因子。
關鍵詞:喬木;光合;大南湖二礦
中圖分類號:Q945
文獻標識碼:A?文章編號:1674-9944(2020)14-0032-03
1?引言
我國是一個以煤炭為主的能源生產和消費大國[1]。開采礦產資源過程中,給經濟帶來快速增長的同時,也不可避免地給生態環境帶來了嚴重的影響,如:地表塌陷、水環境質量破壞、水土流失、沙漠化等[2~5]。哈密地區擁有豐富的煤炭資源,品種多,易開采,在新疆有著舉足輕重的地位。由于大南湖二礦地表主要是原始戈壁,隨著露天煤礦的開采,對地表與地層均有破壞,該礦區植被覆蓋率極低,難以抵抗土壤的侵蝕與搬運,土地沙化、荒漠化加劇[6]。為保障礦區生態環境,需要合理地利用當地鄉土及人工種植植物,科學修復礦區被擾動的環境[7~9]。
光合作用作為植物生長發育的基礎,能夠很大程度地反應出植物對當地的適應狀況[10]。目前,針對荒漠干旱區的植物生理特性的報道較多,如郭洋楠等在新疆克里雅河流域荒漠-綠洲交錯帶研究蘆葦、檉柳、胡楊的光合特性[11]。許疆維等探究荒漠植物-花花柴在高溫脅迫下的生理特性的響應[12]。然而,對礦區這種干旱荒漠區的植被光合生理特性研究較少,本研究選擇榆樹(Ulmus pumila L.)、竹柳(Salix fragilis)、沙棗(Elaeagnus angustifolia L.)3種礦區常用修復植物,這3種植物均具有耐堿、耐干旱的特點。通過探討這3種植物的光合作用日變化以及對環境因子的響應,為礦區進行生態修復和保護提供理論科學依據,同時可對礦區修復植物在該區域的環境脅迫下的生態適應性提供借鑒。
2?材料與方法
2.1?研究區概況
研究區位于哈密市西南部,距哈密市區約84 km。屬于典型的大陸性氣候,年平均氣溫9.8 ℃,極端最高氣溫43 ℃,極端最低氣溫-32 ℃;年降水量33.8 mm,年蒸發量3300 mm,年均日照3358h,無霜期182 d[13]。
2.2?測定方法
在礦區的生態修復示范區中分別選擇榆樹、竹柳、沙棗3種植物,在2019年7月選擇晴朗無云的天氣,從當地時間早上10:00~18:00,每隔2 h采用英國PP-systems公司產的CARAS-2型光合儀分別測定榆樹、竹柳、沙棗葉片的光合參數及環境因子,每株植物選擇3片葉片,植物應選擇選擇健康、完整的葉片,反復測定3次取平均值。光合參數包括:凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)、胞間CO2濃度(Ci);環境因子包括:大氣溫度(Ta)、光合有效輻射(PAR)、大氣相對濕度(RH)、大氣CO2濃度(Ca)。
2.3?數據處理
使用Excel 2016進行數據整理分析及作圖,SPSS 2017進行Pearson相關性分析。
3?結果與分析
3.1?環境因子日變化
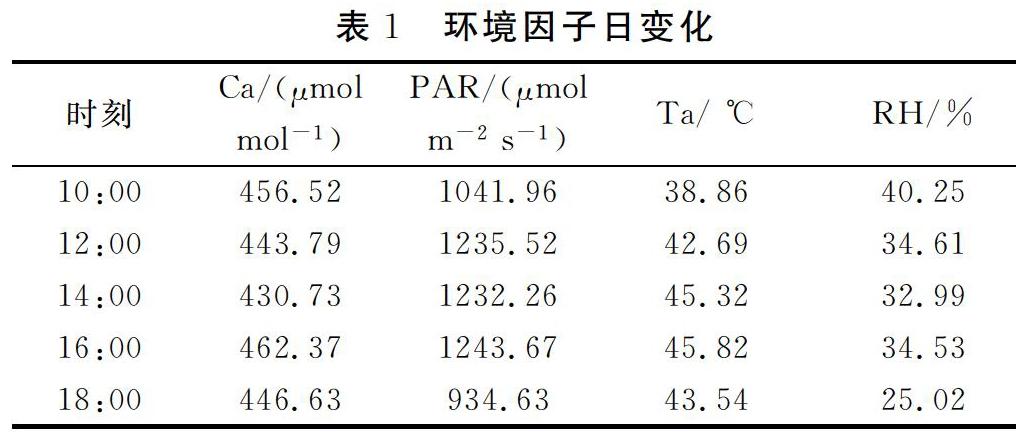
研究區環境因子如表1所示。
由表1可知,礦區在7月的溫度極高,中午可達45 ℃,隨溫度的升高,光合有效輻射增強,二者均先上升再下降;大氣CO2濃度在14:00為最低值;大氣相對濕度早晨最大,下午18:00最小,可能由于礦區熱度具有累積性,在18:00熱度不減小。
3.2?3種植物凈光合速率日變化
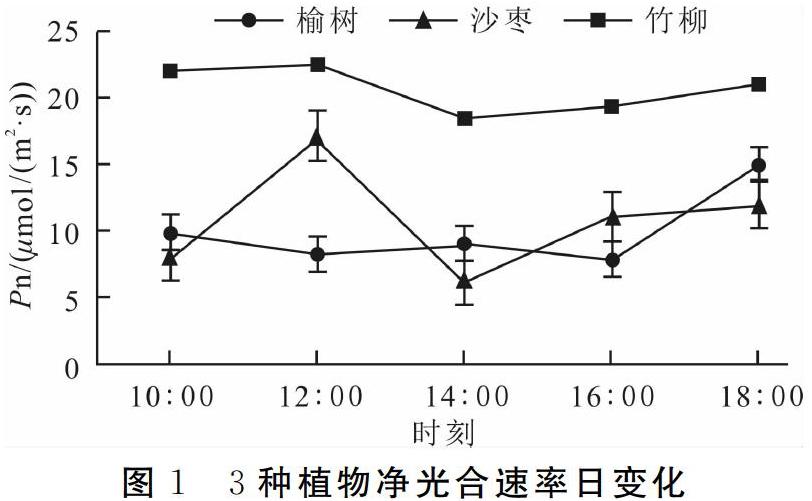
在野外晴朗天氣下,3種植物的凈光合速率日變化如圖1所示。3種植物日均凈光合速率由大到小依次為竹柳(20.61 μmol/(m2·s))>沙棗(10.91 μmol/(m2·s))>榆樹(9.95 μmol/(m2·s))。竹柳的凈光合速率日變化曲線為“V”型,最低值出現在14:00;沙棗的凈光合速率日變化曲線幅度較大,在12:00達到峰值,在14:00為谷值,出現“午休”現象。
3.3?3種植物蒸騰速率日變化
該礦區的3種植物的蒸騰速率日變化如圖2所示,3種植物日均蒸騰速率由大到小依次為沙棗(11.67 mmol/(m2·s))>榆樹(9.41 mmol/(m2·s))>竹柳(8.65 mmol/(m2·s))。3種植物的蒸騰速率日變化與凈光合速率呈相反的趨勢,榆樹從10:00~16:00呈持續上升趨勢,與大氣溫度有相同的趨勢,至16:00到達最高峰(17.63 mmol/(m2·s));竹柳的蒸騰速率日變化呈雙峰型,且第一峰值(12:00)>第二峰值(16:00);沙棗在12:00出現蒸騰“午休”現象,比竹柳出現的“午休”現象提前2 h。
3.4?3種植物氣孔導度日變化
在野外自然狀態下3種植物的氣孔導度日變化如圖3所示,3種植物日均氣孔導度與日均蒸騰速率大小排列一致,由大到小依次為沙棗(235.12 mmol/(m2 s))>榆樹(213.11 mmol/(m2 s))>竹柳(173.44 mmol/(m2 s))。榆樹和沙棗的氣孔導度日變化呈單峰型,沙棗的峰值出現在14:00(305.22 mmol/(m2 s)),榆樹的峰值比沙棗延遲2 h,出現在16:00(352.00 mmol/(m2 s))。竹柳的氣孔導度日變化呈雙峰型,峰值出現在12:00和16:00,且第一峰值(268.00 mmol/(m2 s))>第二峰值(220.33 mmol/(m2 s))。
3.5?3種植物胞間CO2濃度日變化
在大南湖二礦礦區,3種植物的胞間CO2濃度日變化如圖4所示,3種植物日均胞間CO2濃度由大到小依次為沙棗(298.30 μmol/mol)>榆樹(254.70 μmol/mol)>竹柳(191.93 μmol/mol)。整體來看,3種植物在上午的胞間CO2濃度高,日變化曲線為下降的趨勢。榆樹和沙棗在下降過程中在14:00達到峰值。
3.6?3種植物凈光合速率與環境因子的相關性分析
表2為3種植物凈光合速率與環境因子的相關性分析,3種植物中只有沙棗的凈光合速率和蒸騰速率與胞間CO2濃度呈極顯著負相關,相關系數分別為0.667和0.681。3種植物與大氣溫度和大氣CO2濃度分別呈負相關和正相關,但不具有顯著水平;3種植物中僅有沙棗與大氣相對濕度和氣孔導度呈負相關,其余2種植物與大氣相對濕度和氣孔導度呈正相關。
4?討論
植物進行光合作用是其生理活動之一,光合作用日變化反應植物一天中有機物的積累,其動態變化不僅僅與周圍的環境(光照、溫度、水分等)有關,更與自身的生理特性(氣孔、RuBP羧化等)相關[14~16]。本研究對大南湖礦區7月的大氣溫度、相對濕度、光照強度、大氣CO2濃度以及3種植物的光合參數進行測定發現上午隨著光照強度以及溫度的升高,沙棗在12:00達到一天中的最高值,這可能是由于各種環境因子同時作用造成的。之后隨著光照強度以及溫度繼續上升,但大氣相對濕度在降低,3種植物的光合在14:00有了下降的趨勢,這可能是環境因子對其產生了脅迫作用。根據Farquhar的觀點,植物的光合作用下降的原因包括氣孔因素(植物葉片氣孔關閉)和非氣孔因素(葉肉細胞活性降低),由植物凈光合速率與環境因子的相關性可知,榆樹的凈光合速率與蒸騰速率呈負相關,與氣孔導度呈正相關,因此榆樹出現光合“午休”現象是由于非氣孔因素;沙棗是由于自身生理特性及氣孔因素同時作用導致的;竹柳時由于氣孔因素占主導作用[17,18]。在本次研究中,3種礦區植物凈光合速率日均值表現為:竹柳>沙棗>榆樹,從植物種類來看,表明竹柳的光合能力最強,較在礦區生存[3]。
5?結論
(1)在野外自然條件下,7月的3種植物凈光合速率日均值大小表現為:竹柳>沙棗>榆樹,竹柳表現出有較強的光合作用。
(2)影響榆樹、竹柳、沙棗出現光合“午休”現象的原因分別為非氣孔因素、共同作用、氣孔因素。
(3)3種植物日均蒸騰速率由大到小依次為沙棗>榆樹>竹柳。
(4)蒸騰速率與胞間CO2濃度是影響植物凈光合速率的主要環境因子。
參考文獻:
[1]尚海濤.煤炭作為我國基礎能源的重要地位不可動搖[J].中國煤炭,2001,27(4):5~8,13.
[2]卞正富.煤礦區土地復墾條件分區研究[J].中國礦業大學學報,1999(3):37~42.
[3]郭洋楠,孫安安,呂?凱,等.三種強旱生植物在鄂爾多斯礦區生物修復中的生理生態適應性[J/OL].水土保持通報:1-8[2020-06-04].https://doi.org/10.13961/j.cnki.stbctb.20200521.001.
[4]韓克勇,張建亮.礦山開采沉陷災害防治與水環境保護研究[J].太原科技,2008(5):51~52.
[5]張家琦,王?焱,任正興,等.礦區綠色生態修復植物配置模式探索-以陜西省神木市大柳塔煤礦區為例[J].國家林業和草原局管理干部學院學報,2019,18(3):28~32.
[6]Hossain M. Anawar, Farjana Akter, Zakaria M. Solaiman. et al. Biochar: An Emerging Panacea for Remediation of Soil Contaminants from Mining, Industry and Sewage Wastes[J]. Pedosphere,2015,25(5):654~665.
[7]羅春首. 將樂縣礦產集中開采區的礦山地質環境調查與研究[D].福州:福建農林大學,2017.
[8]郭蔚麗,石改新.淺析欒川露采礦山地質環境保護與恢復治理[J].資源導刊,2014(5):10~11.
[9]郭渠江.新疆維吾爾自治區礦山環境生態化法治研究[D].西安:長安大學,2014.
[10]馮大蘭,黃小輝,劉?蕓,等.4種木本植物在石漠化地區的生長狀況及光合特性[J].北京林業大學學報,2015,37(5):62~69.
[11]丁俊祥,鄒?杰,唐立松,等.克里雅河流域荒漠-綠洲交錯帶3種不同生活型植物的光合特性[J].生態學報,2015,35(3):733~741.
[12]許疆維,孫萬豪,吳文博,等.高溫-干旱對沙漠植物花花柴光合作用的影響[J].分子植物育種,2019,17(12):4089~4095.
[13]袁振文.基于新疆哈密氣候特點下的哈密吐哈油田礦區園林綠化研究[J].綠色科技,2012(12):148~149.
[14]裴?斌,張光燦,張淑勇,等.土壤干旱脅迫對沙棘葉片光合作用和抗氧化酶活性的影響[J].生態學報,2013,33(5):1386~1396.
[15]余叔文,湯章城.植物生理與分子生物學[M].2版.北京:科學出版社,1998:155~276.
[16]孫安安,智穎飆,姜平平,等.西鄂爾多斯4種荒漠植物光合作用特征與差異性[J].生態學報,2019,39(13):4944~4952.
[17]Farquhar GD,Sharkey T D.Stomata conductance and photosynthesis[J]. Annual Review of Plant Physiology,1982,33:317~345.
[18]宮江平,高?波,努爾塔依·鐵利汗,等.榆樹秋季衰老葉光合特性研究[J].新疆師范大學學報(自然科學版),2015,34(3):22~28.
