雙香豆素通過抑制PDK1活性增加人膠質瘤細胞對替莫唑胺敏感性的研究
2020-12-25 02:45:48董敬蓉丁文斌
中國實驗診斷學 2020年12期
周 磊,熊 壯,馬 躍,董敬蓉,丁文斌
(長春中醫藥大學附屬醫院,吉林 長春130021)
神經膠質瘤是中樞神經系統最常見原發性腫瘤。由于惡性膠質瘤患者多為青壯年,且缺乏有效的綜合治療措施,因此預后很差。替莫唑胺作為基礎化療藥物廣泛應用于惡性膠質瘤臨床治療中[1]。替莫唑胺是一種新型烷化劑,通過細胞毒作用發揮抗腫瘤效果,較傳統細胞毒藥物不良反應小,患者耐受性好。限制替莫唑胺治療效果的主要原因在于細胞毒藥物的劑量限制性毒性,因此如何增加腫瘤細胞對化療藥物的敏感性一直是抗腫瘤化療研究的重點和難點。
惡性腫瘤細胞特征性的代謝模式為有氧糖酵解,也稱為Warburg效應,而逆轉Warburg效應導致腫瘤細胞代謝重編程,是重要的抗腫瘤治療靶點之一。丙酮酸脫氫酶激酶1(PDK1)可通過磷酸化負調節丙酮酸脫氫酶復合物(PDC)活性,進而影響腫瘤細胞能量代謝[2]。很多研究發現,PDK1在多種惡性腫瘤中高表達,且與腫瘤細胞的侵襲、耐藥、惡性進展中密切相關。雙香豆素作為臨床用藥,具有抗凝、抗腫瘤和抗菌的多重藥效學作用。且已有研究表明,雙香豆素具有抑制PDK1活性的作用[3]。因此,本研究以人膠質瘤U87細胞為模型,探討雙香豆素是否通過抑制PDK1活性進而影響腫瘤細胞代謝模式,以提高U87細胞對替莫唑胺的敏感性。
1 材料與方法
1.1 細胞
人腦星形膠質母細胞瘤U87MG細胞來自吉林大學基礎醫學院病理生理學系實驗室。U87MG細胞常規培養條件培養于含有10%FBS(Hyclone,美國)的DMEM培養基(Gibco,美國)中,置于37℃、5% CO2培養箱。
1.2 細胞存活率實驗(Cell Viability Assays)
U87MG細胞接種于96孔板中,8 000細胞/孔。不同濃度替莫唑胺作用24 h后,每孔加入20 μl MTT(5 mg/ml)孵育4 h后,棄去培養液,加入150 μl DMSO,室溫振蕩 10 min,全自動酶標儀(波長 490 nm)測定各孔 A值,計算細胞存活率。
1.3 免疫印跡
冷RIPA裂解液裂解細胞。全細胞裂解液使用Bio-Rad蛋白試劑測定蛋白濃度。每樣本取約50 μg總蛋白進行SDS-PAGE電泳分離,濕轉法到PVDF膜上,封閉后按照1∶1 000加入特異性一抗,4℃孵育過夜,洗脫后加入HRP標記的二抗,ECL發光法檢測目的蛋白表達情況。
1.4 葡萄糖攝取和乳酸生成檢測
細胞加藥處理后,PBS清洗后,加入新鮮培養液。收集培養液,分別用葡萄糖和乳酸檢測試劑盒(碧云天生物技術有限公司)檢測培養液中葡萄糖和乳酸濃度,用細胞總蛋白定量。
1.5 統計學分析
實驗重復3次,數據使用平均值±標準誤(mean±SE)表示。使用one-way ANOVA進行統計學檢驗,分析軟件為SPSS 12.0,P<0.01有統計學意義。
2 結果
2.1 雙香豆素與替莫唑胺對U87MG細胞存活率的影響
根據文獻及本研究組的前期工作,采用雙香豆素濃度為25 μmol/L,分別聯合替莫唑胺濃度為100 μmol/L、200 μmol/L和400 μmol/L作用于U87MG細胞24 h,MTT法檢測細胞存活率。結果顯示,雙香豆素與替莫唑胺聯合作用對U87MG細胞存活率抑制明顯高于單獨替莫唑胺作用組,特別是替莫唑胺濃度為200 μmol/L時,與雙香豆素合加具有明顯的協同作用(圖1,P<0.01)。所以后續實驗中選用替莫唑胺濃度為200 μmol/L,雙香豆素濃度為25 μmol/L。

*P<0.01圖1 雙香豆素與替莫唑胺對U87MG細胞存活率的影響
2.2 雙香豆素與替莫唑胺對U87MG細胞PDK1活性的影響
PDK1是葡萄糖代謝途徑中的關鍵酶,在糖酵解中發揮重要作用。PDK1通過磷酸化PDH的ser232位點,降低PDH活性,因而抑制丙酮酸的脫羧氧化生成乙酰輔酶A進入線粒體的過程,促進Warburg效應和腫瘤的生長。免疫印跡法檢測PDK1靶蛋白PDH及磷酸化PDH的ser232位點(p-PDHE1A)的表達變化,結果顯示與對照組相比,TMZ組p-PDHE1A無明顯變化,但DIC組和TMZ+DIC組p-PDHE1A表達明顯降低(圖2)。

圖2 雙香豆素與替莫唑胺對U87MG細胞PDK1活性的影響
2.3 雙香豆素與替莫唑胺對U87MG細胞葡萄糖攝取的影響
為進一步探討雙香豆素影響U87MG細胞對替莫唑胺敏感性的機制,我們檢測了葡糖糖代謝相關指標葡萄糖攝取和乳酸生產量。葡萄糖攝取結果顯示,與對照組相比,TMZ組葡萄糖攝取增高,而DIC組葡萄糖攝取降低,特別是TMZ+DIC組與TMZ組相比,葡萄糖攝取明顯降低(圖3,P<0.01)。
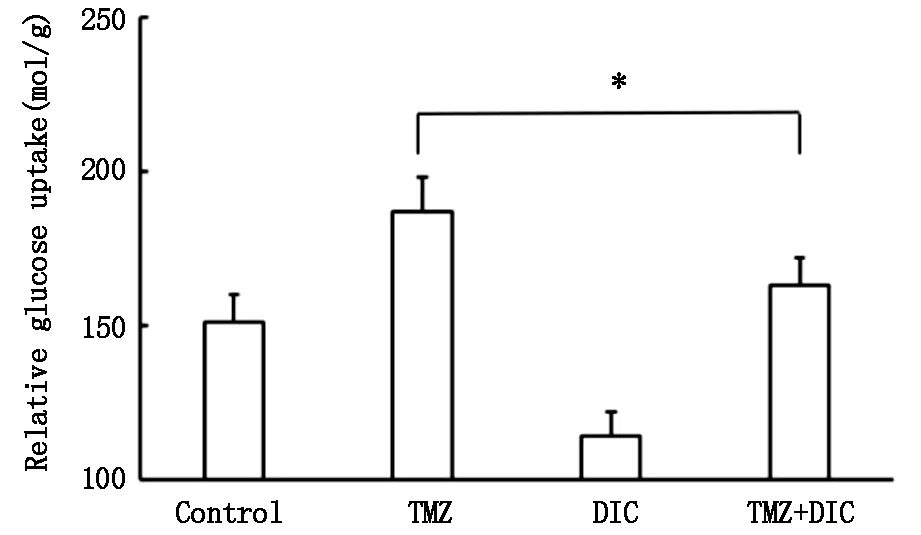
*P<0.01圖3 雙香豆素與替莫唑胺對U87MG細胞葡萄糖攝取的影響
2.4 雙香豆素與替莫唑胺對U87MG細胞乳酸生成的影響
乳酸生成結果顯示,與對照組相比,TMZ組乳酸生成增高,而DIC組乳酸生成降低,特別是TMZ+DIC組與TMZ組相比,乳酸生成明顯降低(圖4,P<0.01)。
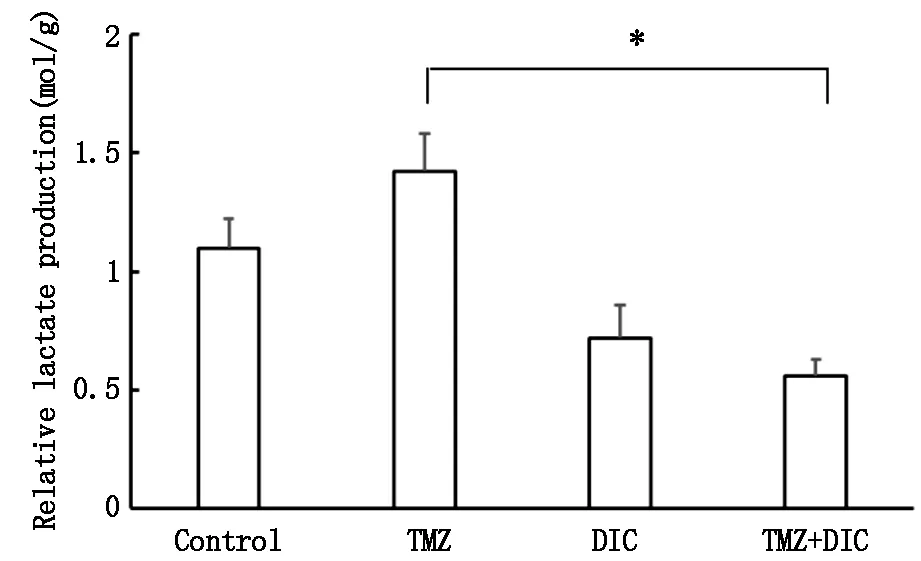
*P<0.01圖4 雙香豆素與替莫唑胺對U87MG細胞乳酸生成的影響
3 討論
惡性膠質瘤預后不佳,特別是化療抵抗是其難以治愈的根本原因之一。惡性膠質瘤細胞可通過致癌信號的激活及不同代謝機制異常導致其對化療治療敏感性降低[4]。神經膠質瘤能量代謝依賴于Warburg效應,即有氧糖酵解。PDK1可通過磷酸化負向調節丙酮酸脫氫酶復合物(PDC),導致PDC失活促進Warburg效應。最新研究證實,PDK1表達與活性受癌基因調節,在多種癌癥中過表達,并在癌細胞代謝中發揮重要作用。
PDK1在腫瘤組織中的高表達與腫瘤中異常糖酵解密切相關,因此,抑制PDK1可作為抗腫瘤治療的新靶點。關于PDKs抑制劑的研究較多,但因具有抗癌療效時劑量過大或者選擇性抑制較差等造成毒副作用較大等問題,限制其在臨床上的應用[5]。Zhou等利用Discovery Studio 3.5軟件的Libdock模塊虛擬篩選并分析了雙香豆素與PDK1的可能結合位點,并在一系列細胞內外實驗中驗證了雙香豆素抑制PDK1活性的效果[6]。此外,Zhang和Xu等在體內外實驗中進一步驗證了雙香豆素可通過抑制PDK1活性進而達到抗卵巢癌作用和增加肝癌細胞化療敏感性的作用[3,7]。
膠質母細胞瘤中PDK1高表達,且其磷酸化水平與患者的不良預后呈正相關。因此,本研究以人膠質瘤U87MG細胞為模型,探討雙香豆素是否可以提高U87MG細胞對替莫唑胺的敏感性。基于文獻報道與本研究組的前期工作,我們選用雙香豆素濃度為25 μmol/L,經免疫印跡實驗證實該濃度對U87MG細胞中PDK1的活性具有很好的抑制效果。而雙香豆素與替莫唑胺聯合作用可明顯增加U87MG細胞對替莫唑胺的敏感性。為進一步探討雙香豆素如何發揮化療增敏作用,我們利用免疫印跡法檢測PDK1靶蛋白PDH及磷酸化PDH的ser232位點(p-PDHE1A)的表達變化,表明雙香豆素單獨或與替莫唑胺聯合作用于U87MG細胞均可有效抑制PDK1活性,而替莫唑胺單獨作用無明顯效果。PDK1是葡萄糖代謝途徑中的關鍵酶,在糖酵解中發揮重要作用。PDK1通過磷酸化PDH的ser232位點,降低PDH活性,因而抑制丙酮酸的脫羧氧化生成乙酰輔酶A進入線粒體的過程,促進Warburg效應和腫瘤的生長。我們在此基礎上,檢測了葡糖糖代謝相關指標葡萄糖攝取和乳酸生產量。替莫唑胺單獨作用可引起U87MG細胞葡萄糖攝取和乳酸生成增多,提示Warburg效應增強可能是腫瘤細胞對抗替莫唑胺的保護性機制之一。而雙香豆素可明顯降低降低替莫唑胺引起的葡萄糖攝取和乳酸生成增多的效應。
綜上,我們的研究結果表明,雙香豆素可通過抑制PDK1活性,改變U87MG細胞代謝模式,進而增加U87MG細胞對替莫唑胺的敏感性。本研究再次驗證了雙香豆素是有效的PDK1活性抑制劑,為其“老藥新用”和臨床惡性膠質瘤綜合治療提供了新的參考。
