網袋埋藏和自然環境下測定森林凋落物早期分解過程的比較
2020-12-25 05:05:38谷永建李玉梅陶千冶
浙江林業科技 2020年6期
谷永建,李玉梅,陶千冶,旺 羅
(1.中國科學院大學地球與行星科學學院,北京 100049;2.中國科學院地質與地球物理研究所,北京 100029;3.中國科學院大學測試中心分子化石實驗室,北京 100049)
作為地球系統碳循環的重要過程,凋落物分解是全球氣候變化研究的熱點之一[1-4]。凋落物分解的研究方法主要有網袋法[5]、小容器法[6]和同位素法[7]等。自20 世紀60 年代起,網袋法在凋落物分解研究中得到廣泛使用[8-10]。該法將凋落物稱量后放入分解袋中,置于土壤表層和地表凋落物之間,定期取回網袋[11],“追蹤”凋落物分解進程和養分釋放過程。網袋法的推廣和使用,能動態監視土壤動物和微生物的群落變化[12],大大促進了凋落物分解過程的定量化研究[13-14],還可為優化生態環境和人工造林提供決策依據[15]。
然而,由于網袋的存在,網袋微環境與自然環境的分解速率存在差異:網袋本身會限制大型土壤動物的進出,從而降低土壤動物對凋落物分解的作用[12];網袋內部可能形成一個相對于外部環境有所差異的微環境,使得凋落物分解過程發生改變[16],小孔徑的網袋造成的微環境可能導致較高的分解率[17]。目前,網袋法作為模擬自然環境中凋落物分解的方法在森林生態研究中應用普遍[11,14],但其與真實自然環境中凋落物分解的比較案例相對較少[18],網袋對凋落物分解的影響尚不十分明確。
分別測定網袋樣品和自然樣品在分解過程的失重率是直接、有效地獲知二者差異的方法[10,13]。但自然凋落物分布在整個凋落區,且可能因風和降水過程等發生移動。若使用失重法,需要將研究區內某一分解時間的所有凋落物進行稱量,工作量大,可操作性差。有必要選擇一種簡單、高效、易操作、能有效指示凋落物分解程度的對比指標。
凋落物的顏色受控于各種色素物質(如葉綠素、類胡蘿卜素、花青素等)及其相對含量[19-20]。在分解作用早期,凋落物的顏色變化明顯,色素濃度不斷下降[21]。通過對植物顏色的定量化研究,紅度(a*)、黃度(b*)和亮度(L*)三種顏色指標與色素含量的關系已經比較明確:
(1)a*:與葉綠素含量負相關,與花色素苷含量正相關[22]。葉片成熟凋落時,葉綠素大部分已經分解[23],對凋落物顏色影響較小,而花色素苷對顏色影響較大。a*指示了花色素苷的含量。
(2)b*:與類胡蘿卜素等黃色色素有關。類胡蘿卜素含量穩定,降解速率低,在葉片衰老時取代葉綠素,導致凋落物呈黃色[19,23-24]。b*指示了類胡蘿卜素等色素的含量。
(3)L*:在凋落物的分解過程中,有機質含量下降、無機成分增加[25]、腐殖質逐漸形成和積累[26],亮度變弱,L*減小。L*直接反映了有機質的分解程度,可作為腐殖質相對含量的指標。
因此,a*,b*和L*作為定量化的顏色值,能夠準確描述凋落物在分解過程中的顏色,進而表征凋落物的分解程度[27],其中,a*,b*和L*分別指示了凋落物花色素苷、類胡蘿卜素和總有機質的分解情況。
基于上述考慮,本文嘗試以a*,b*和L*為基本指標,通過分析北京東靈山暖溫帶落葉闊葉林新鮮凋落物早期分解過程中顏色的變化,考察網袋埋藏和自然埋藏狀態下凋落物的分解速率,以期認識網袋埋藏對凋落物早期分解過程的影響,為定量研究凋落物的分解過程、精確探知地球系統碳循環服務。
1 材料和方法
1.1 研究區概況
東靈山(39°48′~ 40°00′ N,115°24′~ 115°36′ E)位于北京市門頭溝區,距北京城區122 km,屬太行山系,為小五臺山向東南延伸的支脈,主峰海拔2 303 m,是北京市最高峰。該地區屬于暖溫帶大陸季風性氣候區,四季分明,但春、秋短,夏、冬長,冬季寒冷多風且干燥,夏季溫熱多雨,秋季晴朗少風,冷暖適中但雨量偏少。年均氣溫在5~ 11°C,7 月最熱,平均氣溫達到18~ 25°C;1 月最冷,平均氣溫在-10~ -4°C;3-4 月氣溫急劇上升,10-11 月驟然下降,全年無霜期為195 d。年降水量為500~ 650 mm,多集中在夏季,6-8 月的降雨量約占全年降水總量的74%,春、秋季降水量分別占10%和14%,冬季降水量最少,約占2%。地帶性典型植被為暖溫帶落葉闊葉林,以落葉櫟類為主。土壤類型主要為棕色森林土、褐色土和亞高山草甸土[28]。
研究區位于北京東靈山一次生林東坡,海拔在1 500~ 1 600 m,區內主要植被有白樺Betula platyphylla,遼東櫟Quercus wu taishanica和蒙古櫟Q.mongolicus等,有少量色木槭Acer mono和胡桃楸Juglans ma ndshurica伴生。
研究區海拔較高,人跡罕至,有利于試驗的長時間進行,避免了人類活動的干擾。
1.2 樣品采集和處理
1.2.1 自然樣品 在研究區選取100 m×100 m 的永久固定樣地作為1 號樣地。將凋落物自上到下分為F,R,H,S 共四層,依次為當年的新鮮凋落物、半破碎的近年凋落物、破碎程度較高的陳年凋落物和土壤。各層區分明顯,其中,F 層即新鮮凋落物層為本次試驗的研究對象。
于2007 年11 月至2008 年11 月,定期采集凋落物樣品。為保證樣品具有代表性,采用五點取樣法,即:每次采樣時隨機設置A,B,C,D,E 五個采樣點,每個采樣點在<1 m × 1 m 范圍內分別采集3 份F 層凋落物,等量混合后作為該點的樣品。將五個點的樣品等比例高度混合,裝入樣品袋并立即封口,用保溫箱帶回實驗室,在4°C 冰箱內暫時保存備用。
于2007 年初冬、植物全部落葉后的11 月24 日首次采樣。當日F 層凋落物的物理性狀如下:新鮮或輕微變色的未分解凋落物,呈完整片狀,顏色枯黃,干燥易碎,是當年脫落的新鮮凋落物。
為方便度量,以此日為本研究的時間起點,開始計算分解時間。于凋落物分解的第67 天、第133 天、第201 天和第323 天,即2008 年1 月30 日、2008 年4 月4 日、2008 年6 月11 日和2008 年10 月11 日分別用同樣的方法采集F 層樣品。
2008 年夏、秋季節采集樣品時,肉眼觀察并篩選出新凋落物,用鑷子剔除,以免新凋落物輸入影響顏色測定結果。
1.2.2 網袋樣品 選取與上述樣地緊鄰的兩塊100 m×100 m 樣地分別作為2 號樣地和3 號樣地。3 塊樣地的植被類型、地理環境高度一致。
2007 年11 月24 日,用多點網格采樣法收集3 號樣地的F 層凋落物,混合均勻后分裝入尼龍網袋(26 cm×15 cm,孔徑1.1 mm × 0.8 mm)中,每袋裝F 層凋落物20 g。
將裝有凋落物的網袋隨機埋入2 號樣地中的新鮮凋落物下,使網袋微環境與自然環境初始值盡可能保持相同。為方便對照,后續采集自然狀態F 層凋落物的同時,每次隨機選取2~ 5 個網袋,取回網袋樣品,裝入樣品袋中帶回實驗室,在4℃冰箱內暫時保存備用。
此后,于2009 年11 月、2010 年11 月、2013 年4 月、2014 年11 月和2018 年4 月對網袋樣品進行了采集,同樣每次隨機選取2~ 5 個網袋。于2018 年4 月同時采集網袋周邊自然環境下土壤表層凋落物樣品。王瑾等應用網袋法對東靈山不同植物的凋落物分解時間模擬得出平均周轉期為11.64 年[29-30],因此,基本可以判斷土壤表層凋落物至此時已分解了10 年左右,為網袋樣品同期凋落物,可以與網袋埋藏樣品進行對比。
1.3 凋落物顏色測定
凋落物顏色由日本KONICA MINOLTA 公司生產的CM-600d 分光測色計測量,使用L*a*b*色空間。L*a*b*色空間是目前最普遍使用的色空間之一,其中L*表示亮度,在0(黑)~ 100(白)之間變化;a*表示紅度,+a*為紅色方向(0~ 60),-a*為綠色方向(-60~ 0);b*表示黃度,+b*為黃色方向(0~ 60),-b*為藍色方向(-60~ 0)。測試參數:CIE D65 光源(色溫為6 504 K 的正常日光),10°觀察視角。
樣品在70°C 下烘干1 h 后分別粉碎,對測色計進行系統校正后,在保證背景光源恒定的情況下,選取凋落物較為平整的區域手動測量30 次,儀器自動得出測量平均值。測量精度為0.01,誤差為±2%。
凋落物分解過程中顏色不斷發生變化,因此本研究試通過定量顏色值的變化來指示凋落物分解。新鮮凋落物的a*,b*和L*較高,分解一段時間后凋落物的a*,b*和L*降低。
1.4 數據處理方法
數據分析和圖表繪制使用SPSS 22.0 和OriginPro 2015 完成。對不同分解時間的自然樣品和網袋樣品的a*,b*和L*分別進行對數擬合和指數擬合,得到各自擬合回歸方程;同時對凋落物分解早期的三個顏色指標進行Pearson 相關分析,得到相關系數矩陣。
2 結果與分析
2.1 自然環境中凋落物的顏色變化
在自然埋藏環境中,地表凋落物不同取樣時間的顏色變化如圖1。由圖1 可知,a*,b*和L*三個顏色指標均隨著時間的推移而減小,其中,以L*的下降幅度較大,其次是b*,a*的下降幅度較小。分解第1 年,a*的變化范圍為5.51~ 8.84,b*為11.95~ 21.81,L*為32.17~ 50.87。分解10 年后,樣品的a*,b*和L*分別為4.94,10.99 和36.69。

圖1 網袋樣品和自然樣品凋落物顏色值及其變化擬合曲線Figure 1 a*,b* and L* values of samples from litter bag and natural environment and their fitting curves
在10 年的分解期內,對自然環境中的樣品顏色變化的回歸分析顯示,對數擬合曲線更符合凋落物樣品的顏色變化趨勢,其擬合結果如下:

式中,t為分解時間(d),下同。
凋落物顏色的變化主要發生在分解的第1 年(圖1)。在分解早期的第1 年內,盡管a*,b*和L*略有波動,但在整個研究期內凋落物的顏色變化速率相對穩定。凋落物的顏色在第1 年中更趨向于指數變化。進一步分析中,對自然環境中凋落物的a*,b*和L*三個顏色指標隨分解時間的變化進行指數擬合,分別得到:

由指數擬合公式可以看出,a*,b*和L*與分解時間均呈極顯著的負相關。
2.2 網袋微環境中凋落物的顏色變化
將網袋中不同時間凋落物的顏色值在自然環境凋落物的顏色變化趨勢圖上投點,發現網袋中凋落物在分解早期的顏色值雖然分布比較離散,高低值波動較大,但與自然狀態下的凋落物顏色變化趨勢一致。網袋中凋落物的顏色整體上仍表現為隨時間的增加逐漸下降的趨勢(圖1)。在分解的第1 年,a*的變化范圍為6.04~ 8.34,b*為13.86~ 22.02,L*為36.09~ 50.87。分解10 年后,其網袋樣品的顏色值分別下降到4.43,8.74 和31.23。同樣對10 年分解期的網袋樣品顏色值進行了對數擬合:

對分解第1 年的網袋凋落物顏色值分別進行指數擬合,得到分解趨勢線。網袋凋落物顏色值的擬合公式如下:
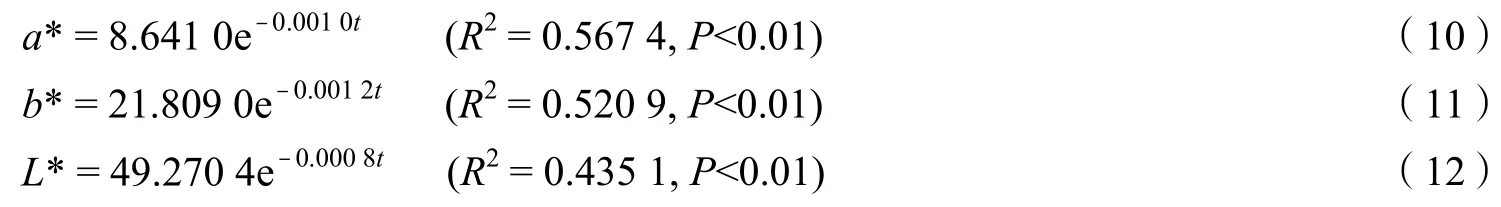
由第1 年的擬合結果顯示,a*,b*,L*觀測值與分解時間之間的相關性均達極顯著水平,且a*與分解時間之間的相關系數 >b*與分解時間之間的相關系數 >L*與分解時間之間的相關系數。
2.3 自然環境和網袋微環境凋落物分解早期顏色變化對比
分解早期,由Pearson 相關分析結果顯示,無論是網袋樣品還是自然環境樣品,其a*,b*,L*三個顏色指標之間呈極顯著的正相關,表明同組樣品顏色變化具有一致性和同步性,即凋落物的L*高時,其a*和b*也高(表1)。L*與a*,b*的顯著性相關表明,凋落物色素的分解趨勢與總有機質的分解趨勢相一致。因此,在未來的工作中,可以考慮用凋落物L*的變化模式代表總體顏色的變化以使問題簡化。

表1 網袋樣品和自然樣品a*,b*,L*的相關分析Table 1 Correlation analysis on a*, b* and L* of samples from litter bag and natural environment
網袋微環境凋落物顏色值主要分布在自然環境凋落物分解擬合曲線兩側,與自然環境凋落物顏色值變化趨勢一致,隨著時間推移而逐漸減小。分解初期,網袋微環境和自然環境中的凋落物顏色值較為接近,隨著分解時間的增加,兩者的差異逐漸增大(圖1),網袋微環境中的凋落物色素變化速率逐漸低于自然環境中的。
網袋中凋落物的a*在分解過程中高于同期自然環境凋落物的a*,表明花色素苷分解速率較低。網袋中凋落物的b*的高值指示類胡蘿卜素等黃色色素分解較慢。網袋中凋落物的L*的高值則說明網袋微環境中凋落物有機質總體分解速率較低。a*,b*和L*三個顏色指標都指示分解早期網袋凋落物的有機質(色素)分解速率要低于自然環境中的。網袋埋藏可能導致凋落物分解速率降低。
2.4 自然環境和網袋微環境凋落物分解后期的顏色變化對比
經過10 年的分解,自然樣品的L*大于分解1 年后的L*。L*主要反映了腐殖質的相對含量[26],由于本研究中利用了土壤表層的凋落物樣品代表自然環境下分解了10 年的凋落物,樣品貼近土壤可能導致土壤的混入使得腐殖質相對含量下降,從而使L*增加,因此測得土壤表層的凋落物樣品的L*高于樣品分解1 年后的L*。對表層土壤的顏色測定發現,土壤的顏色值除a*(4.71)略小于10 年分解后的自然樣品的a*外,b*(11.73)和L*(38.99)均明顯高于自然樣品。可以推斷自然樣品的實際L*應低于32.17,a*和b*也同樣低于測定值。
從10 年的分解趨勢線上可以看出,自然樣品和網袋樣品的顏色值在第1 年迅速下降,在之后9 年的時間里緩慢下降。自然樣品的L*在1 年后已經下降到32.17,只比經過10 年分解后的網袋樣品的L*(31.23)略高,a*和b*的變化與之相同。由于網袋對凋落物分解的抑制作用,在分解過程中,自然樣品的顏色值可能先達到相對穩定值,網袋樣品的顏色值逐漸向這一穩定值靠近,最終達到與自然樣品同樣的顏色值。因此,經過10 年的分解后,自然樣品的實際顏色值與網袋樣品的顏色值相近,或可能低于網袋樣品的顏色值。
3 討論
凋落物分解的影響因素主要包括凋落物基質成分、氣候、微生物和土壤動物等[1-2,12,31],其中,植被的固有特性即凋落物基質成分是主控因素,不同類型的植被凋落物分解速率差異明顯[5]。本研究中,網袋和自然環境中的凋落物成分基本相同,可將其排除。因此,物理化學條件、微生物以及土壤動物可能是影響網袋和自然環境中的凋落物色素分解速率差異的主要因素,具體包括濕度、土壤動物和微生物等。
3.1 濕度
濕度(降水)是影響凋落物分解的重要氣候因子[32]。宋新章等對中國東部氣候帶凋落物分解特征的研究結果表明,降水量是影響中國東部凋落物分解速率的首要氣候因子[33]。凋落物的分解主要發生在夏季雨水多發期。降水一方面通過淋溶作用,造成可溶有機質和易分解碳水化合物的淋失和分解,降水量越大,表層凋落物的解體就越快[34]。另一方面,降水通過影響微生物的活動和酶活性間接影響凋落物的分解[32,34]。
與自然樣品相比,網袋的保護作用導致袋內凋落物結構穩定,降水的直接淋溶和破碎效果降低[35],水溶性物質的淋失強度下降,凋落物的解體速率減緩,色素的分解速率下降。花青素作為一類水溶性植物色素[36],降水有助于其直接從凋落物中流失,而網袋對降水的阻礙使得水溶性色素的流失速率降低,a*下降變慢。其他水溶性色素的降解速率也存在不同程度的下降。
降水影響土壤濕度,而濕度直接影響土壤動物的活性。一定的濕度能夠顯著增加生物多樣性和豐富群落組成[37],水分不足使分解者種群數量下降、活性降低[38]。由于水分不易進入網袋造成內部濕度變低,微生物可利用水資源不足,活性下降,對色素等有機質分解效率減緩,不利于凋落物的分解,使網袋樣品的分解速率低于自然環境樣品,導致顏色變化速率降低,表現為a*,b*,L*變化較慢。
3.2 土壤動物和微生物
土壤生物在分解早期的作用小于氣候,但也是至關重要的[34]。土壤動物(螞蟻、蚯蚓等)主要對凋落物進行破碎,使凋落物表面積增大,屬于物理性分解,破碎產生的凋落物碎屑容易被分解者利用;色素等有機質在分解者(主要為真菌、細菌)及各種酶系統作用下發生生物降解,復雜的有機化合物(色素)被分解為簡單的小分子和腐殖質,使凋落物顏色發生變化,屬于化學分解[11,14,17,25]。土壤動物的破碎盡管只在很小程度上改變凋落物的顏色,但形成的小顆粒有助于土壤微生物的進一步分解。土壤生物在凋落物色素變化中扮演重要角色。微生物活動和酶活性是影響凋落物有機質和色素分解的最直接和關鍵因子,微生物活動強則色素分解速率快[25]。
分解開始階段,土壤動物能夠進入網袋對凋落物進行破碎,微生物作用較小[11],網袋樣品和自然樣品色素分解差異小表現不明顯。隨著時間的推移,網孔的阻礙作用逐漸積累,并且網孔可能被外界物質所覆蓋或者網孔變小,部分土壤動物不能進入網袋,凋落物的破碎作用減弱,使得微生物對小顆粒凋落物的可利用效率下降。同時網孔的遮蔽或變小,使得與外界的營養交換產生變化,氧氣不充足。網孔越小,通氣性愈差。營養元素和氧氣的缺乏導致凋落物微生物活性降低,群落功能多樣性下降[34]。凋落物為土壤動物和微生物提供能量,群落結構越復雜,活性越高,則需要消耗更多能量和養分,對凋落物的分解越快。網袋阻礙土壤動物進入以及對微生物活動的減弱都使得凋落物的消耗下降,從而使網袋樣品色素分解慢于自然樣品,顏色變化差距逐漸增大。
總之,網袋的存在既削弱了降水對凋落物的淋溶破碎,同時導致袋內濕度降低,在分解過程中孔徑變小又阻止了土壤動物的進入,使凋落物的破碎速度下降,小顆粒有機質供應不足。濕度不足和與外界交換的不暢通使得微生物活性和數量受到影響,在一定程度上限制了微生物活性和降低了酶活性,對凋落物色素等有機質的分解變慢[35],造成網袋凋落物顏色相對于自然環境變化較小。由于網袋在凋落物早期分解過程中的抑制作用,根據網袋凋落物早期分解過程預估的周轉期可能要大于凋落物的實際周轉期[29]。
另外,網袋將袋內和袋外兩種環境分開,分解初期氣候條件和生物作用相同,隨著時間的延長,不同網袋逐漸形成各自內部的微環境,使得各自的分解模式產生差異[35],從而導致凋落物顏色與時間的相關性較差(圖1)。在外部環境相同的情況下,微環境的影響不容忽視[17]。不同網袋內逐漸形成相對穩定的微環境,使得網袋內溫度和濕度發生改變,生物作用產生變化,較高溫度和濕度導致較高的分解速率[16]。微環境的不同造成色素分解差異明顯,這可能是導致本次研究中網袋微環境中凋落物顏色值分布離散、規律性差的原因。
4 結論
凋落物分解過程中,顏色指標a*,b*,L*隨著時間的增加逐漸降低,三者顯著正相關。a*,b*,L*可以分別指示凋落物花色素苷、類胡蘿卜素和總有機質的分解速率。
分解早期,網袋微環境與自然環境中凋落物的顏色值差異小,隨分解時間的增加差異逐漸增大。總體上,網袋樣品的色素分解速率低于自然環境樣品的。這可能是網袋的存在阻礙了降水淋溶、降低網袋內樣品濕度、阻礙了土壤動物的活動及抑制土壤微生物的繁殖等因素造成的。經過1 年的分解之后,凋落物的顏色值變化減慢,顏色逐漸趨于穩定。
作為野外測定森林凋落物分解速率的常用方法,網袋對凋落物的分解具有一定的抑制作用,在利用網袋進行凋落物分解研究時應考慮其影響。
