河南省首次發現梨孢鐮刀菌引起的小麥赤霉病
2020-12-28 02:23:48徐飛王俊美楊共強
植物保護 2020年6期
徐飛 王俊美 楊共強


摘要 2015年從河南省田間小麥赤霉病病穗上分離得到一種生長速度較慢的鐮刀菌,通過形態學和分子鑒定明確其分類地位,通過田間單小花滴注法和噴霧法接種測定其致病力,并通過高效液相色譜串聯質譜分析對麥穗中的毒素種類進行測定,明確其產毒特征。結果表明:分離得到的8個菌株均為梨孢鐮刀菌,在馬鈴薯葡萄糖瓊脂(PDA)培養基上為白色菌落,菌落底部產生少量紅色色素,平均生長速度為13.3 mm/d;小型分生孢子為橢球形葡萄狀,平均大小為7.1 μm×5.8 μm,未見大型分生孢子和厚垣孢子;致病力弱,且不侵染穗軸,單小花滴注法接種條件下平均病級為0.1,噴霧法接種條件下平均病小穗率為6.5%;供試的8個鐮刀菌菌株均不產生T-2和HT-2毒素,均產生雪腐鐮刀菌烯醇(NIV)毒素,NIV毒素含量水平為371.74~5 282.80 μg/kg,其中3個菌株產生少量脫氧雪腐鐮刀菌烯醇(DON)毒素(86.13~227.22 μg/kg)。
關鍵詞 小麥; 小麥赤霉病; 梨孢鐮刀菌; 雪腐鐮刀菌烯醇
中圖分類號: S 435.121.45
文獻標識碼: A
DOI: 10.16688/j.zwbh.2019483
Abstract In 2015, eight slow-growing Fusarium strains were isolated from the diseased wheat heads with Fusarium head blight in the field in Henan province. The species status was identified by morphological and molecular characters. The pathogenicity of these isolates was determined by the single-spikelet point inoculation and spray inoculation, and the toxin types of these isolates in wheat heads were determined by HPLC/electrospray ionization tandem mass spectrometry system. The results showed that all the eight isolates were Fusarium poae, with white colonies on PDA and a small amount of red pigments produced at the bottom of the colony. The average growth rate of these isolates was 13.3 mm/d. The average size of microconidia was 7.1 μm×5.8 μm, globose and napiform, without macroconidia and chlamydospores. The pathogenicity of these isolates was weak, and they did not infect the spike axis. The average disease level was 0.1 under the single-spikelet point inoculation. The average spikelet rate was 6.5% under the spray inoculation. These isolates did not produce T-2 and HT-2, but all of them produced nivalenol (NIV). The level of NIV was 371.74-5 282.80 μg/kg, and three isolates produced a small amount of deoxynivalenol (DON) (86.13-227.22 μg/kg).
Key words wheat; Fusarium head blight; Fusarium poae; nivalenol
由鐮刀菌引起的小麥赤霉病不僅能造成產量損失,而且病原菌產生毒素污染籽粒,造成人畜中毒[1]。禾谷鐮刀菌復合群Fusarium graminearum species complex,黃色鐮刀菌F.culmorum和燕麥鐮刀菌F.avenaceum是分布廣泛的主要致病種,而梨孢鐮刀菌F.poae也是引起小麥赤霉病的重要病原之一[2]。梨孢鐮刀菌不僅能產生B型非大環類單端孢霉烯族類毒素,包括雪腐鐮刀菌烯醇(nivalenol,NIV)、脫氧雪腐鐮刀菌烯醇(deoxynivalenol,DON)、3-乙酰基脫氧雪腐鐮刀菌烯醇(3-acetyldeoxynivalenol,3ADON)及15-乙酰基脫氧雪腐鐮刀菌烯醇(15-acetyldeoxynivalenol,15ADON),還能產生A型非大環類單端孢霉烯族類毒素,包括T-2毒素和HT-2毒素[3]。由于梨孢鐮刀菌產毒種類多,且在不同地域或氣候條件下產生不同的毒素[3],而我國梨孢鐮刀菌產毒情況未見報道,因此我國的梨孢鐮刀菌的產毒情況仍需研究。
近年來,梨孢鐮刀菌在阿根廷、加拿大、德國和意大利等國家的小麥上分離頻率增加[4-5]。在我國僅湖北報道4個菌株為梨孢鐮刀菌[6],之后未有報道。本研究旨在明確河南省梨孢鐮刀菌的分布和產毒種類。
1 材料與方法
1.1 材料來源
供試鐮刀菌的小麥病穗樣本于2015年采集自河南省新鄉市獲嘉縣(35°3′45″N, 113°37′51″E, 海拔82.8 m和35°3′44″N, 113°37′51″E, 海拔79.5 m)和濮陽市濮陽縣(35°32′3″N, 115°13′54″E, 海拔53.1 m)的小麥-水稻連作地塊。
1.2 病原菌的分離純化和形態學鑒定
將樣品中發病的小穗剪下,剪掉麥芒,在超凈工作臺里分別在70%乙醇中處理10 s,3%NaClO中消毒1 min,然后用無菌水沖洗3次;用滅菌的濾紙片吸干水分并在工作臺中吹干;最后將病組織置于含有抗生素的馬鈴薯葡萄糖瓊脂培養基(PDA,150 μg/mL硫酸鏈霉素和 75 μg/mL氨芐青霉素鈉鹽)上,于25℃培養箱中黑暗條件下培養3~7 d,待病組織周圍長出類似鐮刀菌菌落后,挑取至新的PDA培養基上培養。每個發病的麥穗分離1個菌株。隨后連續3次挑取邊緣菌絲進行純化獲得純化菌株。將分離得到8個菌株的菌絲塊分別放入20%甘油-75℃凍存。
將收集來的康乃馨葉片制作康乃馨葉片培養基[7],制作方法為將葉片沖洗干凈后,再用無菌水沖洗,剪成約10 mm×10 mm的小塊,放入錫箔紙中包裹,在70℃條件下烘干3 h。在直徑6 cm的培養皿中倒入滅菌的2%水瓊脂培養基,凝固后用無菌的鑷子將6塊烘干的康乃馨葉片放在培養基上。將各純化菌株挑取至康乃馨葉片培養基中間,在25℃條件下,12 h黑暗12 h光照條件下培養7~14 d。挑取葉片邊緣的分生孢子進行顯微觀察,測量并記錄其大小。每個菌株記錄20個孢子并計算其平均值。比對鐮刀菌鑒定手冊中的孢子大小進行初步鑒定[7]。
1.3 分子鑒定
將各菌株用挑針轉移到PDA培養基上,25℃培養7 d后,用無菌牙簽挑取氣生菌絲50~100 mg裝在2 mL的離心管中。然后用液氮速凍,存放在-20℃的冰箱里待用。每個2 mL的離心管加入5顆直徑2 mm的鋼珠(提前加入試劑盒中提取液300 μL),使用德國萊馳球磨儀(MM400)30次/s振蕩2 min,然后加入試劑盒中提取液200 μL,使用E.Z.N.A.TM HP Fungal DNA Kit (OMEGA, 貨號:D3195)按提取步驟進行DNA提取。DNA提取后,置于1.5 mL的離心管中于-20℃保存備用。擴增8個菌株的EF-1α基因序列進行系統發育分析,確定其種類,PCR引物和程序參考ODonnell等[8]和Proctor等[9]。引物EF1 (5′-ATGGGTAAGGA(A/G)GACAAGAC-3′)和EF2 (5′-GGA(G/A)GTACCAGT(G/C)ATCATGTT-3′)用來擴增EF-1α基因部分序列,擴增產物送至生工生物工程(上海)股份有限公司測序。然后使用特異性引物Fp82F(5′-CAAGCAAACAGGCTCTTCACC-3′)和 Fp82R(5′-TGTTCCACCTCAGTGACAGGTT-3′)進行特異性檢測,進一步明確其種類[10]。
1.4 致病力測定
將-75℃凍存的鐮刀菌菌株在新鮮的PDA平板上活化,25℃黑暗條件下培養3 d,打取5塊(5 mm)邊緣菌絲塊轉接到裝有羧甲基纖維素酯培養基(CMC)(carboxymethyl-cellulose 150 g/L, NH4NO3 1.0 g/L, KH2PO4 1.0 g/L, MgSO4·7H2O 0.5 g/L, 酵母提取物1.0 g/L)的小三角瓶(100 mL),25℃,150 r/min搖培3~7 d,待產生大量分生孢子后,分別將供試菌株的分生孢子懸浮液經擦鏡紙過濾,用血球計數板對濾液中分生孢子進行計數,用無菌水調整濃度為1×107個/mL和1×106個/mL分別用于單小花滴注接種和噴霧接種。禾谷鐮刀菌對照菌株15HJ1-12用相同的方法獲得分生孢子,其單小花滴注接種的分生孢子濃度為5×105個/mL。
于小麥品種‘周麥18揚花初期(2016年5月3日),采用單小花滴注法和噴霧法兩種方法分別接種。單小花滴注法采用剪刀剪去麥穗中部小穗的內外穎殼頂部少許,滴入鐮刀菌分生孢子懸浮液20 μL,每個麥穗接種1個小穗,每個品種接種30穗,10穗為1個重復,分別套透明塑料袋保濕,并記錄接種后的天氣情況,2 d后去掉保濕袋,接種后第10 天分別記錄每穗的病級,病級標準參照《小麥抗病蟲性評價技術規范 第4部分:小麥抗赤霉病評價技術規范》(NY/T 1443.4-2007)[11]中單小花滴注法的嚴重度分級來進行調查記載,平均病級=調查穗總病級/總調查穗數;噴霧法接種采用鐮刀菌分生孢子懸浮液噴霧,每平方米噴霧50 mL,每個品種接種30穗,10穗為1個重復,分別套透明塑料袋保濕,接種后第10天分別記錄每穗的病小穗數和總小穗數,并計算病小穗率,接種后10 d的病小穗率=病小穗數/總小穗數×100%。小麥成熟后,將噴霧法接種的病穗分別裝入網袋,晾干后4℃冰箱中保存,以進行后續的麥穗組織毒素種類和含量測定。
1.5 毒素種類測定
將收回的麥穗剪碎,然后使用德國萊馳粉碎儀(ZM200, 1.5 mm不銹鋼環篩)進行粉碎,過20目篩裝入不同的自封袋中備用。每個樣品粉碎后,嚴格清理以避免相互污染。寄往江蘇省農業科學院農產品質量安全與營養研究所通過高效液相色譜串聯質譜分析進行產毒類型和產生量分析[12]。
2 結果與分析
2.1 病原菌形態學鑒定
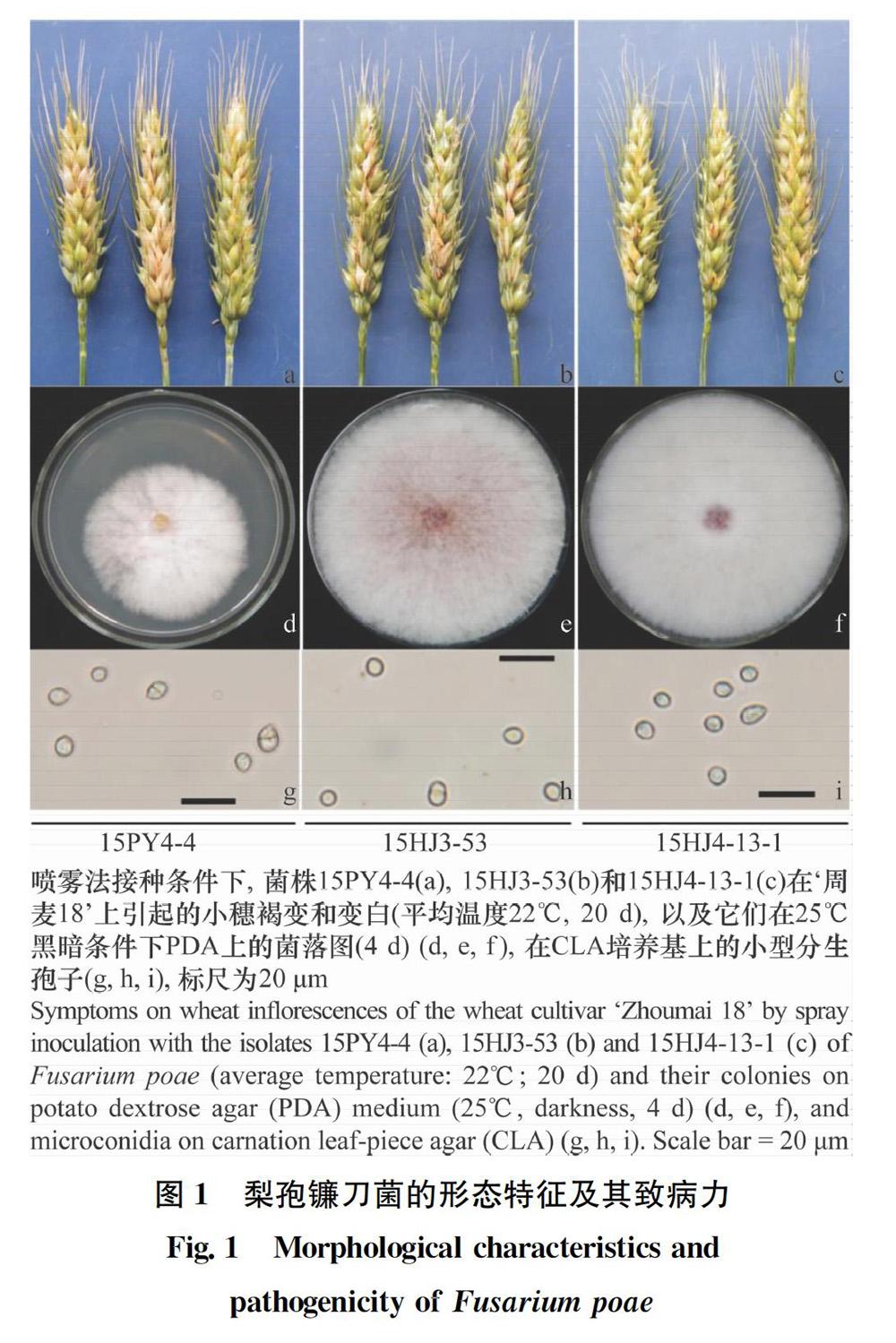
在25℃黑暗條件下,8個菌株在PDA上菌落為白色,底部能產生少量紅色色素,生長速度為12.0~14.3 mm/d(平均為13.3 mm/d);其小型分生孢子呈橢球形,葡萄狀,多數無分隔,偶見1個分隔,大小為(6.3~7.9)μm×(5.4~6.2)μm(平均為7.1 μm×5.8 μm),未見大型分生孢子和厚垣孢子(圖1,表1)。形態學鑒定結果與梨孢鐮刀菌相符。
2.2 分子鑒定
通過EF-1α基因序列測定,整合后在GenBank上進行BLAST比對,結果表明:8個菌株的序列(登錄號為:KX298968~KX298972, KX298974~KX298976)與DNA序列庫中登錄號為MG889924和KP639705的梨孢鐮刀菌的EF-1α基因序列同源性為100%。通過特異性引物Fp82F/R擴增,電泳后均產生0.22 kb的條帶,再次確認8個菌株為梨孢鐮刀菌。
2.3 致病力測定和再分離結果
供試的8個鐮刀菌菌株接種后保濕2 d的條件下,單小花滴注法接種10 d后,70%~90%的接種小穗不發病,10%~30%的接種小穗能夠產生明顯褐變,且不侵染至穗軸,8個鐮刀菌菌株的平均病級為0.1~0.3(平均為0.1)。禾谷鐮刀菌對照菌株15HJ1-12的平均病級為1.2。噴霧法接種10 d后,小穗能產生明顯褐變,但不侵染穗軸,病小穗率為1.6%~15.8%(平均為6.5%)。將噴霧法接種后的發病小穗按照菌株分別取樣,每個菌株3個小穗,進行組織分離,分離得到白色菌落菌株,并用裝50 mL CMC進行搖培,得到橢圓形葡萄狀小型分生孢子,因此確定為梨孢鐮刀菌(圖1,表1)。
2.4 毒素種類測定
供試的8個鐮刀菌菌株均不產生T-2和HT-2毒素,均產生NIV毒素,NIV毒素含量水平為371.74~5 282.80 μg/kg。另外,8個菌株中,只有菌株15PY4-4產生ZEN毒素(20.12 μg/kg);4個菌株不產生DON、3ADON和15ADON毒素;3個菌株(15PY4-4、15HJ4-49和15HJ4-13-1)產生DON毒素(86.13~227.22 μg/kg),2個菌株(15HJ4-13-1和15PY4-4)產生3ADON毒素(58.64、159.23 μg/kg),3個菌株(15PY4-4、15HJ3-53和15HJ4-13-1)產生15ADON毒素(29.11~293.14 μg/kg)(表1)。
3 結論與討論
河南省小麥赤霉病菌主要是禾谷鐮刀菌和亞洲鐮刀菌[13],而梨孢鐮刀菌較少。單小花滴注接種條件下,梨孢鐮刀菌(平均病級為0.1)致病力比禾谷鐮刀菌(平均病級為1.2)弱,且不侵染穗軸。梨孢鐮刀菌致病力極弱,可能是其沒有在中國大面積發生的原因。但是Audenaert等[14]發現三唑類殺菌劑(戊唑醇、丙硫菌唑、氟環唑和葉菌唑)防治小麥赤霉病造成梨孢鐮刀菌的比例上升。隨著我國小麥赤霉病菌對多菌靈抗性的升高,戊唑醇已經成為防治小麥赤霉病的主要藥劑,并且丙硫菌唑及其復配藥劑已在中國登記并使用。因此監測梨孢鐮刀菌在中國的流行對我國小麥赤霉病的防治具有重要意義。
河南省梨孢鐮刀菌主要分布在新鄉市獲嘉縣的紅旗渠和濮陽市濮陽縣的沿黃稻-麥連作田塊中,可能原因是梨孢鐮刀菌更喜歡濕潤的環境[15]。河南省梨孢鐮刀菌主要產生雪腐鐮刀菌烯醇毒素,不產生T-2和HT-2毒素,與阿根廷菌株一樣[5],而與挪威菌株產生HT-2毒素不一樣[16]。本研究首次在河南省發現梨孢鐮刀菌,其主要分布在沿黃稻麥連作區,致病力弱并主要產生雪腐鐮刀菌烯醇毒素,為梨孢鐮刀菌的監測流行提供有利信息。
參考文獻
[1] BAI G, SHANER G. Scab of wheat: prospects for control [J]. Plant Disease, 1994, 78(8): 760-766.
[2] 陸維忠, 程順和, 王裕中. 小麥赤霉病研究[M]. 北京: 科學出版社, 2001.
[3] STENGLEIN S A. Fusarium poae: a pathogen that needs more attention [J]. Journal of Plant Pathology, 2009, 91(1): 25-36.
[4] XU Xiangming, PARRY D, NICHOLSON P, et al. Predominance and association of pathogenic fungi causing Fusarium ear blight in wheat in four European countries [J]. European Journal of Plant Pathology, 2005, 112(2): 143-154.
[5] STENGLEIN S A, DINOLFO M I, BARROS G, et al. Fusarium poae pathogenicity and mycotoxin accumulation on selected wheat and barley genotypes at a single location in Argentina [J]. Plant Disease, 2014, 98(12): 1733-1738.
[6] ZHANG Jingbo, WANG Jianhua, GONG Andong, et al. Natural occurrence of Fusarium head blight, mycotoxins and mycotoxin-producing isolates of Fusarium in commercial fields of wheat in Hubei [J]. Plant Pathology, 2013, 62(1): 92-102.
[7] LESLIE J F, SUMMERELL B A. The Fusarium laboratory manual [M]. Oxford, UK: Blackwell Publishing, 2006.
[8] ODONNELL K, WARD T J, GEISER D M, et al. Genealogical concordance between the mating type locus and seven other nuclear genes supports formal recognition of nine phylogenetically distinct species within the Fusarium graminearum clade [J]. Fungal Genetics and Biology, 2004, 41(6): 600-623.
[9] PROCTOR R H, MCCORMICK S P, ALEXANDER N J, et al. Evidence that a secondary metabolic biosynthetic gene cluster has grown by gene relocation during evolution of the filamentous fungus Fusarium [J]. Molecular Microbiology, 2009, 74(5): 1128-1142.
[10]PARRY D W, NICHOLSON P. Development of a PCR assay to detect Fusarium poae in wheat [J]. Plant Pathology, 1996, 45(2): 383-391.
[11]中華人民共和國農業部. 小麥抗病蟲性評價技術規范—第4部分: 小麥抗赤霉病評價技術規范NY/T 1443.4-2007 [S]. 北京:中國標準出版社,2007:1-6.
[12]QIU Jianbo, SHI Jianrong. Genetic relationships, carbendazim sensitivity and mycotoxin production of the Fusarium graminearum populations from maize, wheat and rice in eastern China [J]. Toxins, 2014, 6(8): 2291-2309.
[13]徐飛, 楊共強, 王俊美, 等. 河南省小麥赤霉病菌種群組成及致病力分化 [J]. 植物病理學報, 2016, 46(3): 294-303.
[14]AUDENAERT K, LANDSCHOOT S, VANHEULE A, et al. Impact of fungicide timing on the composition of the Fusarium head blight disease complex and the presence of deoxynivalenol (DON) in wheat[M]∥NOORUDDIN T. Fungicides: Beneficial and harmful aspects. Rijeka, Croatia: InTech, 2011: 79-98.
[15]LACEY J, BATEMAN G L, MIROCHA C J. Effects of infection time and moisture on development of ear blight and deoxynivalenol production by Fusarium spp. in wheat [J]. Annals of Applied Biology, 1999, 134(3): 277-283.
[16]LANGSETH W, RUNDBERGET T. The occurrence of HT-2 toxin and others trichothecenes in Norwegian cereals [J]. Mycopathologia,1999, 147(3): 157-165.
(責任編輯:田 喆)
