微衛(wèi)星DNA標(biāo)記評(píng)估湛江光裸星蟲養(yǎng)殖群體的遺傳多樣性
2020-12-28 02:42:29姚澤彬姚維彭錦勝王慶恒郭昱嵩
南方農(nóng)業(yè)·上旬 2020年11期
姚澤彬 姚維 彭錦勝 王慶恒 郭昱嵩

摘? ?要? ?為評(píng)估湛江地區(qū)光裸星蟲養(yǎng)殖群體的種質(zhì)現(xiàn)狀,利用11對(duì)熒光標(biāo)記微衛(wèi)星引物對(duì)光裸星蟲的1個(gè)野生群體和2個(gè)養(yǎng)殖群體,每個(gè)群體取30~31個(gè)樣本進(jìn)行遺傳多樣性分析。結(jié)果表明,11 個(gè)微衛(wèi)星位點(diǎn)在不同群體呈現(xiàn)出不同程度的多態(tài)性,野生群體和2個(gè)養(yǎng)殖群體的平均等位基因數(shù)(Na)在4.182~4.636,平均有效等位基因數(shù)(Ne)在2.447~2.632,平均觀測(cè)雜合度(Ho)、期望雜合度(He)分別在0.442~0.471、0.525~0.536,平均多態(tài)信息含量(PIC)為0.461~0.475。Hardy-Weinber 平衡分析顯示,3 個(gè)群體的大部分位點(diǎn)未偏離平衡。在每個(gè)群體中進(jìn)行連鎖不平衡分析,發(fā)現(xiàn)有4對(duì)位點(diǎn)間顯著(P<0.05)或者極顯著(P<0.01)偏離連鎖平衡。分子方差分析(AMOVA)顯示,光裸星蟲大部分的變異來自于群體內(nèi)而非群體間。綜合分析認(rèn)為,本研究采集的2個(gè)光裸星蟲養(yǎng)殖群體遺傳多樣性豐富,暫未出現(xiàn)近交衰退現(xiàn)象。
關(guān)鍵詞? ?光裸星蟲;遺傳多樣性;微衛(wèi)星標(biāo)記
中圖分類號(hào):S932? ?文獻(xiàn)標(biāo)志碼:A? ? DOI:10.19415/j.cnki.1673-890x.2020.31.001
光裸星蟲(Sipunculus nudus L.,1766),隸屬于方格星蟲綱(Sipunculidea)方格星蟲目(Sipunculiformes)方格星蟲科(Sipunculidae)方格星蟲屬(Sipunculus),俗稱“沙蟲”,生長(zhǎng)于溫帶及熱帶水域的潮間帶,以沙粒間的底棲藻類、有機(jī)碎屑等為食,在中國(guó)沿海均有分布。光裸星蟲營(yíng)養(yǎng)成分豐富,味道鮮美,高蛋白、低脂肪,具有較高的食用價(jià)值[1],但自然采捕難度大,且過度采捕、海區(qū)污染、灘涂破壞等威脅光裸星蟲的種質(zhì)資源[2],自然采捕已經(jīng)難以滿足市場(chǎng)的長(zhǎng)期需求。近年,光裸星蟲的人工養(yǎng)殖日益成熟[3-7],然而,養(yǎng)殖業(yè)的快速發(fā)展常會(huì)因無序引種、苗種異地交易等導(dǎo)致種質(zhì)資源混雜,進(jìn)而影響遺傳多樣性的長(zhǎng)期穩(wěn)定。因此,有必要對(duì)人工養(yǎng)殖光裸星蟲的遺傳多樣性進(jìn)行監(jiān)測(cè)和評(píng)估。
光裸星蟲的遺傳多樣性研究目前主要集中在對(duì)分子標(biāo)記和線粒體DNA的應(yīng)用上,王慶恒等運(yùn)用RAPD技術(shù)對(duì)4個(gè)光裸星蟲地理群體的遺傳多樣性進(jìn)行研究,發(fā)現(xiàn)不同地理群體存在較大的遺傳分化[8]。郭昱嵩等利用微衛(wèi)星DNA分子標(biāo)記分析了北部灣3個(gè)光裸星蟲地理群體的遺傳多樣性,得出“群體間存在較大的基因流、已產(chǎn)生較低水平的遺傳分化”的結(jié)論[9]。Du等利用線粒體COⅠ序列分析了3個(gè)地理野生群體的遺傳多樣性差異,認(rèn)為光裸星蟲目前具有較高的遺傳多樣性和獨(dú)特的種群結(jié)構(gòu)[10]。彭銀輝等基于線粒體控制區(qū)序列開展了6個(gè)地理野生群體光裸方格星蟲的遺傳多樣性研究,發(fā)現(xiàn)遺傳分化在群體間并不顯著,光裸方格星蟲具有較高的遺傳多樣性,且不同地理群體存在頻繁的基因交流[11]。周于娜等利用線粒體控制區(qū)序列分析了光裸星蟲不同地理野生群體和不同地理養(yǎng)殖群體的遺傳差異情況,認(rèn)為養(yǎng)殖群體光裸星蟲的遺傳多樣性略低于野生群體,養(yǎng)殖群體正在逐漸積累遺傳變異[12]。
微衛(wèi)星DNA(microsatellite DNA),又稱簡(jiǎn)單序列重復(fù)(SSR,simple sequence repeat),是1~6個(gè)堿基串聯(lián)重復(fù)序列。SSR標(biāo)記具有穩(wěn)定性好、多態(tài)性豐富、引物通用性較好和共顯性遺傳等特點(diǎn),是研究群體遺傳學(xué)的常用手段。目前,運(yùn)用微衛(wèi)星標(biāo)記研究光裸星蟲遺傳多樣性的報(bào)道較少。
本研究運(yùn)用科研團(tuán)隊(duì)自主開發(fā)的SSR標(biāo)記,篩選出多態(tài)性高的標(biāo)記對(duì)光裸星蟲3個(gè)群體的遺傳多樣性進(jìn)行檢測(cè),并以野生群體作為參考群體來評(píng)估養(yǎng)殖光裸星蟲的遺傳多樣性,從而為光裸星蟲種質(zhì)資源保護(hù)和合理利用提供參考依據(jù),還可為養(yǎng)殖光裸星蟲的種苗優(yōu)劣評(píng)估提供基礎(chǔ)數(shù)據(jù)支持。
1 材料與方法
1.1 實(shí)驗(yàn)材料及DNA提取
實(shí)驗(yàn)用野生光裸星蟲采集自湛江遂溪縣下六鎮(zhèn)(遂溪野生群體,SX)的自然灘涂。2個(gè)湛江養(yǎng)殖群體分別采集自湛江碧海灣水產(chǎn)有限公司樂民繁育基地(樂民養(yǎng)殖群體,LM)和廣東海洋大學(xué)覃斗(覃斗養(yǎng)殖群體,TD)試驗(yàn)基地。參照文獻(xiàn)[13]的苯酚/氯仿抽提法,稍作修改后用于提取光裸星蟲基因組DNA,經(jīng)1%的瓊脂糖凝膠電泳檢測(cè)后,用凝膠成像儀進(jìn)行觀察,估算DNA的濃度和純度。
1.2 微衛(wèi)星序列的引物篩選
本實(shí)驗(yàn)中項(xiàng)目組自主開發(fā)20對(duì)光裸星蟲微衛(wèi)星引物[14],委托上海生工生物工程有限公司合成。將合成好的微衛(wèi)星引物進(jìn)行群體初篩選,篩選實(shí)驗(yàn)用20個(gè)隨機(jī)個(gè)體,提取DNA后對(duì)所有引物進(jìn)行預(yù)擴(kuò)增,根據(jù)預(yù)擴(kuò)增的結(jié)果篩選出11對(duì)多態(tài)性引物(見表1)。預(yù)擴(kuò)增體系與程序如下:PCR預(yù)擴(kuò)增總體系為10 μL,包含DNA模板0.5 μL,上下游引物(5 μmol·L-1)各0.5 μL,PCR Mix 5 μL,剩余體積用ddH2O 補(bǔ)齊至10 μL。PCR反應(yīng)程序?yàn)椋?4 ℃預(yù)變性80 s;94 ℃
30 s,退火(溫度依據(jù)引物而定)30 s,72 ℃ 45 s,30個(gè)循環(huán);72 ℃延伸5 min。
PCR產(chǎn)物在8%非變性聚丙烯酰胺凝膠分離,銀染顯色,以出現(xiàn)清晰主帶為標(biāo)準(zhǔn),篩選出多態(tài)性引物和每對(duì)引物最適退火溫度,等位基因大小以 20 bp DNA Ladder Marker為參照標(biāo)準(zhǔn)進(jìn)行判讀。
1.3 PCR擴(kuò)增及微衛(wèi)星檢測(cè)
委托上海生工生物工程有限公司合成熒光引物,用于光裸星蟲3個(gè)群體樣本的擴(kuò)增,每個(gè)群體30~32個(gè)個(gè)體。
PCR擴(kuò)增體系為25 μL,包含DNA模板2 μL,上下游引物(5 μmol·L-1)各1 μL,Taq酶0.125 μL,10×PCR Buffer(含Mg2+) 2.5 μL,dNTPs (2.2 mmol·L-1) 2 μL,剩余體積用ddH2O 補(bǔ)齊至25 μL。每個(gè)微衛(wèi)星標(biāo)記隨機(jī)抽取4個(gè)PCR產(chǎn)物進(jìn)行檢測(cè),用1%瓊脂糖凝膠進(jìn)行電泳,然后放入凝膠成像儀中觀察,檢測(cè)PCR產(chǎn)物是否正常。將檢測(cè)到無異常的PCR產(chǎn)物送往上海生工生物工程有限公司進(jìn)行毛細(xì)管電泳檢測(cè),檢測(cè)儀器為3730XL序列分析儀(ABI公司,美國(guó))。
1.4 數(shù)據(jù)分析
使用GenAlEx 6.503軟件對(duì)生物公司反饋的基因片段長(zhǎng)度等數(shù)據(jù)進(jìn)行遺傳分析,計(jì)算11個(gè)位點(diǎn)的有效等位基因數(shù)(Ne)、等位基因數(shù)(Na) 、期望雜合度(He)、觀測(cè)雜合度(Ho)、Hardy-Weinberg平衡檢驗(yàn)、固定指數(shù)(Fis)、群體間遺傳距離(DA)、遺傳相似系數(shù)(I)和分子方差分析(AMOVA)。利用PIC-CAL0.6軟件計(jì)算每個(gè)微衛(wèi)星位點(diǎn)的多態(tài)信息含量(PIC)。利用PopGen32軟件對(duì)每個(gè)群體的連鎖不平衡進(jìn)行檢測(cè)。無效等位基因用Micro-checker軟件進(jìn)行分析。群體間基于Nei's遺傳距離的聚類樹使用MEGA-X軟件進(jìn)行繪制。
2 結(jié)果與分析
2.1 微衛(wèi)星引物的篩選
篩選實(shí)驗(yàn)結(jié)果顯示,微衛(wèi)星多態(tài)位點(diǎn)有11個(gè),其余9個(gè)為單態(tài)位點(diǎn)。所篩選出來的11對(duì)多態(tài)性引物的特征見表1。
2.2 等位基因數(shù)和有效等位基因數(shù)
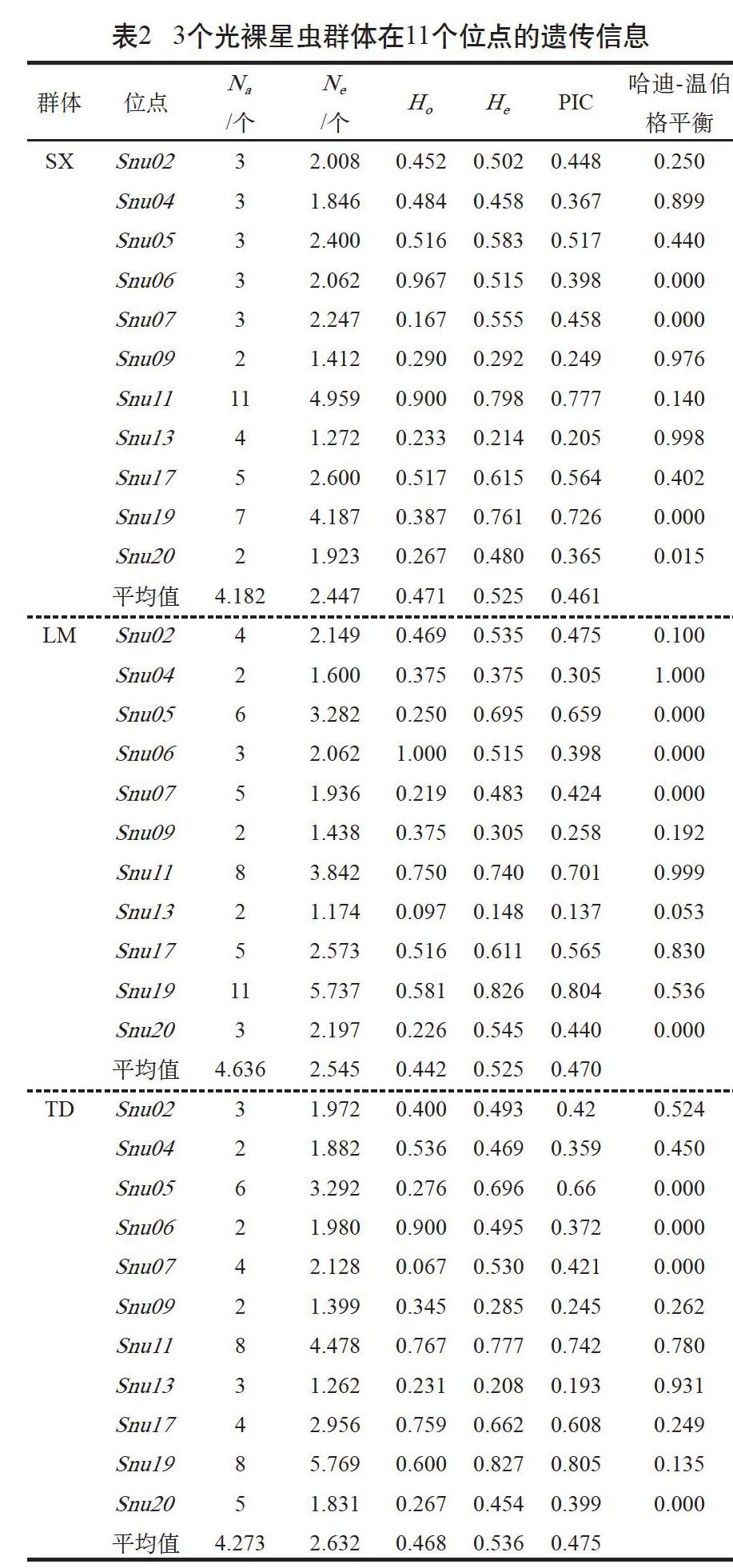
3個(gè)光裸星蟲群體的11個(gè)微衛(wèi)星標(biāo)記均表現(xiàn)出多態(tài)性,由表2可知,3個(gè)群體中,遂溪野生群體的平均Na和平均Ne均最低,為4.182個(gè)和2.447個(gè);樂民養(yǎng)殖群體的平均Na最高,為4.636個(gè);覃斗養(yǎng)殖群體的平均Ne最高,為2.632個(gè)。
2.3 遺傳雜合度、Hardy-Weinberg平衡與連鎖不平衡
表2數(shù)據(jù)顯示,3個(gè)群體的平均觀測(cè)雜合度Ho和期望雜合度He的范圍分別在0.442~0.471和0.525~0.536,其中,平均Ho遂溪野生群體最大,為0.471,樂民養(yǎng)殖群體最小,為0.442;覃斗養(yǎng)殖群體的平均He在3個(gè)群體中最大,為0.536,另外2個(gè)群體的平均He均為0.525。經(jīng)Hardy-Weinberg平衡的卡方檢驗(yàn),在3個(gè)群體11個(gè)多態(tài)微衛(wèi)星位點(diǎn)中大部分未偏離平衡狀態(tài)(P>0.05),每個(gè)群體均有4個(gè)位點(diǎn)偏離平衡狀態(tài)(P<0.05),其中Snu06、Snu07和Snu20位點(diǎn)在3個(gè)群體中均偏離平衡,Snu05位點(diǎn)在2個(gè)養(yǎng)殖群體偏離平衡狀態(tài),Snu19位點(diǎn)在野生群體偏離平衡。在每個(gè)群體中進(jìn)行連鎖不平衡分析,結(jié)果表明4對(duì)位點(diǎn)間顯著(P<0.05)或者極顯著(P<0.01)偏離連鎖平衡;在樂民養(yǎng)殖群體中有2對(duì)連鎖不平衡(Snu05/Snu19、Snu07/Snu13),野生群體和覃斗養(yǎng)殖群體均只出現(xiàn)1對(duì)相同的連鎖不平衡(Snu7/Snu19)。
2.4 多態(tài)信息含量(PIC)與無效等位基因
3個(gè)群體11個(gè)微衛(wèi)星位點(diǎn)的平均PIC在0.461~0.475,遂溪野生群體最小,2個(gè)養(yǎng)殖群體相對(duì)較大。Micro-checker檢測(cè)發(fā)現(xiàn),Snu07、Snu19和Snu20位點(diǎn)在3個(gè)群體中均存在無效等位基因,Snu05位點(diǎn)在2個(gè)養(yǎng)殖群體中存在無效等位基因,其余位點(diǎn)在各個(gè)群體中均不存在無效等位基因。
2.5 群體間遺傳分化分析
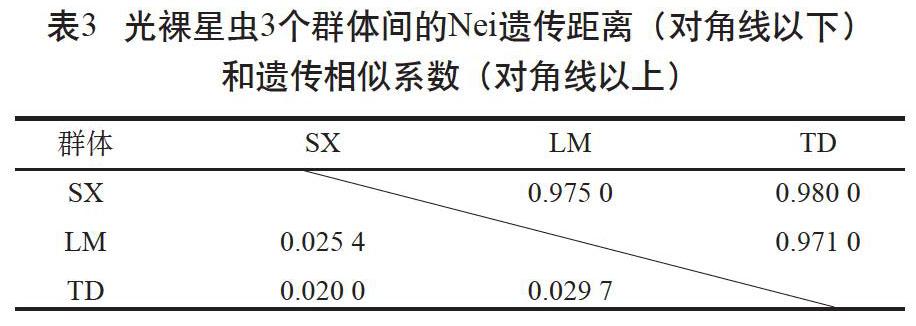
利用GenAlEx 6.503軟件計(jì)算得到了3個(gè)群體間的Nei遺傳距離(DA)和遺傳相似系數(shù)(I)(見表3)。樂民養(yǎng)殖群體和覃斗養(yǎng)殖群體間的DA最大,I最小;遂溪野生群體和覃斗養(yǎng)殖群體間的DA最小,I最大。3個(gè)群體間的DA為0.020~0.034。根據(jù)Nei遺傳距離采用鄰接法和UPGMA 法對(duì)3個(gè)群體進(jìn)行聚類分析,結(jié)果顯示遂溪野生群體和樂民養(yǎng)殖群體聚為一支,覃斗養(yǎng)殖群體為一支(見圖1,僅列出鄰接圖)。
種群的遺傳分化系數(shù)(Fst)表明種群遺傳分化的水平,F(xiàn)st在0~0.05表明種群遺傳分化很弱,F(xiàn)st在0.05~0.15表明種群遺傳分化處于中等水平,F(xiàn)st在0.15~0.25表明種群遺傳分化較大,F(xiàn)st大于0.25表明種群遺傳分化很大。本研究中方格星蟲3個(gè)群體的Fst在0.008~0.012,均不顯著。AMOVA分析表明,99%的遺傳變異源于群體內(nèi),群體間的遺傳變異僅占1%(見表4)。
3 討論
遺傳雜合度(H)是指微衛(wèi)星位點(diǎn)為雜合子的比例,可反映各群體在多個(gè)位點(diǎn)上的遺傳變異,是描述群體遺傳變異的一個(gè)重要指標(biāo)。群體H高,表明該群體的遺傳變異多,群體遺傳多樣性高[15]。郭昱嵩等在2012年報(bào)道了北部灣三個(gè)光裸星蟲野生群體的遺傳多樣性[9],我們用與該研究相同的11個(gè)多態(tài)性位點(diǎn)對(duì)其平均雜合度進(jìn)行矯正,得到湛江烏石群體的平均Ho、平均He分別為0.458、0.500,北海山口群體的平均Ho、平均He分別為0.471、0.484,越南錦普群體的平均Ho、平均He分別為0.484、0.532。矯正后的數(shù)據(jù),本研究遂溪野生群體的平均Ho和平均He略高于郭昱嵩等報(bào)道的湛江烏石野生群體,2個(gè)養(yǎng)殖群體的平均Ho和平均He除了樂民群體的平均Ho低于烏石群體外,其他均略高于烏石群體,但差異很小,這表明光裸星蟲遂溪群體的遺傳變異程度目前仍處于較高水平。
多態(tài)信息含量(PIC)反映了微衛(wèi)星標(biāo)記所包含的遺傳信息含量。在一個(gè)群體中該雜合子的比例越大,PIC值越大,該雜合子提供的遺傳信息越多。根據(jù)Botstein等[16]提出的標(biāo)準(zhǔn),本研究的11個(gè)多態(tài)性位點(diǎn)中有4個(gè)高度多態(tài)性位點(diǎn)(PIC>0.5)、6個(gè)中度多態(tài)性位點(diǎn)(0.25 遺傳距離(DA)的大小可以用來確定群體之間的親緣關(guān)系,群體間的DA與親緣關(guān)系呈正比,與遺傳相似度(I)呈反比。DA越大,表明群體間的親緣關(guān)系越遠(yuǎn),I越小;反之亦然[17]。本研究采用11個(gè)微衛(wèi)星位點(diǎn)對(duì)3個(gè)光裸星蟲群體進(jìn)行遺傳多樣性分析,根據(jù)Nei's指數(shù)法對(duì)光裸星蟲3個(gè)群體的遺傳結(jié)構(gòu)進(jìn)行分析,結(jié)果顯示遂溪野生群體和覃斗養(yǎng)殖群體之間的I最大(0.980)、DA最小(0.020),鄰接法和UPGMA法聚類分析為一支。遺傳分化指數(shù)(Fst)是指群體內(nèi)的變異位點(diǎn)在群體位點(diǎn)中所占的比值,比值越大,群體遺傳分化程度越高,本研究中光裸星蟲3個(gè)群體間的Fst值在0.008~0.012,均小于0.05,3個(gè)群體均分化較小[18],這可能與養(yǎng)殖群體為遂溪野生群體繁育的后代有關(guān)。AMOVA分析表明,群體間的變異僅1%,提示群體內(nèi)的遺傳變異是引起種群總體變異的主要因素。這與光裸星蟲海球幼體時(shí)期浮游生活相吻合,因?yàn)檫@增加了群體間的基因交流,導(dǎo)致群體間的變異程度遠(yuǎn)不如群體內(nèi)部。總的來說,光裸星蟲養(yǎng)殖群體和野生群體之間分化很小,養(yǎng)殖群體暫未出現(xiàn)明顯的退化,這與周于娜等[12]報(bào)道的結(jié)果相似。
整體而言,本研究篩選的11個(gè)微衛(wèi)星位點(diǎn)具有較高的多態(tài)性,適用于3個(gè)光裸星蟲群體的遺傳多樣性研究。研究結(jié)果表明遂溪野生群體與2個(gè)養(yǎng)殖群體的各項(xiàng)遺傳多樣性參數(shù)之間總體上差異不大,群體間分化水平較低,光裸星蟲養(yǎng)殖群體暫未出現(xiàn)明顯的退化現(xiàn)象,這與現(xiàn)階段光裸星蟲大群體親本育苗、沙蟲苗養(yǎng)殖群體沙灘放養(yǎng)等養(yǎng)殖模式吻合。但是,近年來海洋環(huán)境污染特別是海灘垃圾污染導(dǎo)致的灘涂破壞有加重趨勢(shì)[19-21],加上人為無序引種、苗種異地交易等,可能會(huì)進(jìn)一步導(dǎo)致光裸星蟲資源的遺傳多樣性水平下降,因此保護(hù)光裸星蟲種質(zhì)資源已經(jīng)刻不容緩。我們認(rèn)為只有加強(qiáng)對(duì)海洋垃圾源頭的控制,定期檢測(cè)光裸星蟲資源的遺傳多樣性水平,同時(shí)推廣科學(xué)合理的養(yǎng)殖模式,才能保護(hù)好光裸星蟲資源,達(dá)到星蟲資源的可持續(xù)利用。
參考文獻(xiàn):
[1] 羅少杰,楊創(chuàng)業(yè),王慶恒,等.光裸星蟲4個(gè)野生群體的營(yíng)養(yǎng)成分分析與品質(zhì)評(píng)價(jià)[J].廣東海洋大學(xué)學(xué)報(bào),2016,36(1):25-30.
[2] 王紅勇.海南沿海光裸方格星蟲生態(tài)習(xí)性及其資源保護(hù)[J].河北漁業(yè),2014(10):16.
[3] 蔣艷,蔡德建,鄒杰,等.方格星蟲苗種池塘中間培育試驗(yàn)研究[J].廣西科學(xué),2010,17(2):175-177.
[4] 鄒杰,彭慧婧,蔣艷,等.方格星蟲親體培育試驗(yàn)[J].漁業(yè)現(xiàn)代化,2010,37(3):30-33.
[5] 文雪,蔣艷,鄒杰,等.方格星蟲灘涂人工養(yǎng)殖試驗(yàn)[J].科學(xué)養(yǎng)魚,2011(1):38-39.
[6] 陳振國(guó),班庭輝,陳振華,等.方格星蟲土池育苗技術(shù)規(guī)程[J].海洋與漁業(yè),2015(8):64-65.
[7] 王慶恒,鄧岳文,張家煒,等.一種光裸星蟲室內(nèi)苗種培育的方法:CN106035246A[P].2019.
[8] 王慶恒,杜曉東,李康.光裸星蟲遺傳多樣性的RAPD分析[J].海洋水產(chǎn)研究,2006(3):57-61.
[9] 郭昱嵩,王慶恒,黎幸連,等.北部灣光裸星蟲3個(gè)地理群體遺傳多樣性[J].中國(guó)水產(chǎn)科學(xué),2012,19(1):62-69.
[10] Du X, Chen Z, Deng Y, et al. Comparative Analysis of Genetic Diversity and Population Structure of Sipunculus nudus as Revealed by Mitochondrial COI Sequences[J]. Biochemical Genetics, 2009,
47(11-12):884-891.
[11] 彭銀輝,周于娜,劉旭佳,等.基于線粒體控制區(qū)序列的光裸方格星蟲群體遺傳多樣性分析 [J].水產(chǎn)學(xué)報(bào),2017,41(10):1542-1551.
[12] 周于娜,彭銀輝,劉旭佳,等.光裸方格星蟲野生與養(yǎng)殖群體線粒體控制區(qū)序列的遺傳差異分析[J].水生生物學(xué)報(bào),2017,41(2):384-390.
[13] Sambrock J, Russel DW. Molecular Cloning: a Laboratory Manual (3rd Edition)[M]. NewYork:Cold Spring Harbor Laboratory Press, 2002: 461-471.
[14] Wang Qingheng, Guo Yusong, Wang Zhongduo, et al. Isolation and Characterization of Microsatellite DNA Loci From the Peanut Worm, Sipunculus nudus[J]. Genetics and Molecular Research: Gmr, 2012, 11(2): 1662-1665.
[15] 劉楚吾,黎錦明,劉麗,等.中國(guó)龍蝦微衛(wèi)星標(biāo)記的篩選及遺傳多樣性分析[J].遺傳,2010,32(7):737-743.
[16] Botstein D, White RL, Skolnick M, et al. Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J]. Am Jhum Genet, 1980, 32(3): 314-331.
[17] 張愛兵,王正軍,譚聲江,等.分子生態(tài)學(xué)重要概念——遺傳距離及其測(cè)度的理論研究概況[J].生態(tài)學(xué)報(bào),2002,22(6):149-155.
[18] Balloux F, Lugon-moulin N. The Estimation of Population Differentiation with Microsatellite Markers[J]. Molecular Ecology, 2002, 11(2): 155-165.
[19] 黎樹式,林俊良,黃鵠,等.廣西海灘侵蝕原因與修復(fù)[J].北部灣大學(xué)學(xué)報(bào),2019,34(12):30-37.
[20] 莫珍妮,曹慶先,陳圓,等.廣西沿海典型海灘海洋垃圾調(diào)查研究初探[J].化學(xué)工程與裝備,2018(7):299-301.
[21] 趙肖,綦世斌,廖巖,等.我國(guó)海灘垃圾污染現(xiàn)狀及控制對(duì)策[J].環(huán)境科學(xué)研究,2016,29(10):1560-1566.
(責(zé)任編輯:丁志祥)
