用Cyt b 基因分析松嫩平原區(qū)湖泊水庫(kù)大銀魚(yú)的遺傳多樣性
2021-01-06 02:34:58魯翠云陳昕那榮濱鄭先虎李喆唐富江
水產(chǎn)學(xué)雜志 2020年5期
魯翠云,陳昕,那榮濱,鄭先虎,李喆,唐富江
(中國(guó)水產(chǎn)科學(xué)研究院黑龍江水產(chǎn)研究所,黑龍江 哈爾濱 150070)
大銀魚(yú)Protosalanx hyalocranius 隸屬于鮭形目Salmoniformes 銀魚(yú)科Salangidae,自然分布于我國(guó)錢塘江河口、長(zhǎng)江口及長(zhǎng)江下游湖泊、黃海、渤海沿岸河口及朝鮮半島西海岸和越南等。銀魚(yú)類均為一年生的小型魚(yú)類,大銀魚(yú)是銀魚(yú)類群中個(gè)體較大的物種,曾作為重點(diǎn)經(jīng)濟(jì)魚(yú)類在我國(guó)北方大、中型湖泊和水庫(kù)廣泛移植[1-3]。近年來(lái),松嫩平原區(qū)湖泊水庫(kù)大銀魚(yú)增殖漁業(yè)迅速發(fā)展,已成為我國(guó)最大的大銀魚(yú)產(chǎn)區(qū),為漁業(yè)提質(zhì)增效做出了貢獻(xiàn),也對(duì)大銀魚(yú)資源起到了遷地保護(hù)的作用。大銀魚(yú)的研究主要集中在分類、生物學(xué)、移植增殖生態(tài)學(xué)上[4-6],在遺傳學(xué)研究方面開(kāi)展得相對(duì)較少。研究表明大銀魚(yú)的同工酶和隨機(jī)擴(kuò)增多態(tài)性DNA(random amplified polymorphic DNA,RAPD)的多樣性均低于太湖新銀魚(yú)Neosalanx taihuensis 和寡齒新銀魚(yú) Neosalanx oligodontis[7,8]。
線粒體細(xì)胞色素b(Mitochondrial cytochrome b,Cyt b)基因進(jìn)化速率快,約為16S rRNA 的四倍[9],適合于近緣物種間的系統(tǒng)進(jìn)化研究。用Cyt b 基因序列研究銀魚(yú)科魚(yú)類的系統(tǒng)分化較多。羅宏偉等[10]研究表明三峽庫(kù)區(qū)大銀魚(yú)Cyt b 基因序列多態(tài)性較低。李大命等[11,12]對(duì)大銀魚(yú)Cyt b 基因分析表明太湖和洪澤湖群體均為單倍型多樣性高而核苷酸多樣性低的種群,即經(jīng)過(guò)強(qiáng)捕撈遺傳瓶頸效應(yīng)后伴隨著迅速的種群增長(zhǎng)與突變的積累而形成。Xiao 等[13]指出淮河大銀魚(yú)線粒體Cyt b 基因多樣性較高。對(duì)于移植到北方的大銀魚(yú)種群,Tang 等[14]分析了移植到黑龍江、松花江、興凱湖等大銀魚(yú)的遺傳分化,結(jié)果顯示移植群體的遺傳多樣性較原種地太湖有大幅增加。在此基礎(chǔ)上,本研究利用Cyt b 全序列分析了移植到北方松嫩平原區(qū)湖泊、水庫(kù)的大銀魚(yú)群體的遺傳分化,可為大銀魚(yú)種群的遺傳管理和持續(xù)利用提供支持。
1 材料與方法
1.1 材料
大銀魚(yú)采自松嫩平原區(qū)的5 個(gè)湖泊水庫(kù),分別為黑龍江省大慶市連環(huán)湖(LHH)72 尾和龍虎泡(LHP)49 尾,吉林省松原市查干湖(CGH)54 尾、長(zhǎng)春市新立城水庫(kù)(XLC)22 尾和四平市二龍山水庫(kù)(ELS)29 尾,共226 尾。分別取其尾鰭,無(wú)水乙醇固定后,-20℃冷凍保存?zhèn)溆谩?/p>
1.2 基因組DNA 提取
采用傳統(tǒng)的酚氯仿抽提法從大銀魚(yú)鰭條組織中提取基因組DNA[15],使用NanoDropTM8000 分光光度計(jì)檢測(cè)所提取DNA 的濃度及純度,稀釋至50 ng/μL,4℃保存?zhèn)溆谩?/p>
1.3 PCR 擴(kuò)增與測(cè)序
使用引物L(fēng)14321 和H15634 擴(kuò)增大銀魚(yú)Cyt b基因全序列[16],L14321 引物序列為5’-CCAGTGA CTTGAAAAACCACCG-3’;H15634 引物序列為5’-CTTAGCTTTGGGAGTTAAGGGT-3’,引物由上海生工生物公司合成。PCR 反應(yīng)體系為25 μL,其中模板DNA 2 μL、混合PCR 緩沖液buffer 18 μL(10 mmol/L Tris-Cl(pH8.0)、50 mmol/L KCl、1.5 mmol/L MgCl2、200 μmol/L dNTP)、上下游引物(10 μmol/L)各0.5 μL、Taq DNA 聚合酶1.5 U,其余體積用去離子水補(bǔ)充。擴(kuò)增反應(yīng)在ABI 9700 型PCR 儀上完成,反應(yīng)程序?yàn)椋?4℃預(yù)變性7 min;94℃變性30 s,56℃退火45 s,72℃延伸1 min,30 個(gè)循環(huán);72℃延伸5 min。PCR 產(chǎn)物經(jīng)2%瓊脂糖凝膠電泳檢測(cè)后,由上海生工生物工程技術(shù)服務(wù)有限公司純化后進(jìn)行雙向測(cè)序及拼接。
1.4 序列分析
測(cè)序后,將序列輸入Clustal X 軟件[17]進(jìn)行序列的對(duì)位排列,并加以人工校對(duì),截取相同長(zhǎng)度的序列用于群體遺傳分析。用DnaSP v5 軟件[18]統(tǒng)計(jì)變異位點(diǎn)類型和數(shù)目、計(jì)算單倍型數(shù)和核苷酸多態(tài)性等;用MEGA 7.0 軟件[19]分析序列的堿基組成和差異百分比、變異位點(diǎn)、簡(jiǎn)約信息位點(diǎn)數(shù)、轉(zhuǎn)換/顛換值,用Kimura 2-Parameters 方法計(jì)算5 個(gè)群體間的遺傳距離,分別構(gòu)建基于鄰接法(Neighbor-Joining,NJ)和非加權(quán)配對(duì)算術(shù)平均法(unweighted pair group method using arithmetic average,UPGMA)的群體間發(fā)生關(guān)系的聚類樹(shù),采用Bootstrap(重復(fù)數(shù)=1 000)檢驗(yàn)分子系統(tǒng)樹(shù)各分支的置信度。利用Arlequin 3.11軟件[20]中的分子方差分析(AMOVA)計(jì)算群體間的遺傳分化系數(shù)(Fst)及遺傳變異組成。
2 結(jié)果與分析
2.1 核苷酸和氨基酸序列的特征
將所得序列在GenBank 中進(jìn)行Blast 比對(duì),結(jié)果確定為大銀魚(yú)線粒體Cyt b 基因的全序列[21]。所有序列經(jīng)Clustal X 比對(duì)并加以人工校對(duì)后,截取1 141 bp 序列用于群體遺傳分析。經(jīng)MEGA 7.0 分析,此序列可編碼380 個(gè)氨基酸。保守位點(diǎn)1 081個(gè),變異位點(diǎn)60 個(gè),其中單變異位點(diǎn)32 個(gè),簡(jiǎn)約信息位點(diǎn)28 個(gè)。測(cè)得的序列中A、T、C、G 的堿基組成分別為21.73%、29.27%、32.33%和16.67%,其中A+T 含量(51%)略高于C+G 含量(49%),轉(zhuǎn)換/顛換值為3.808。
2.2 單倍型及核苷酸多樣性
5 個(gè)群體226 個(gè)樣本的Cyt b 基因可分為38 個(gè)單倍型,其中單倍型Hap 4 的個(gè)體數(shù)最多(31.42%),其次為單倍型Hap 7(22.57%),且Hap 4和Hap 7 的個(gè)體分布于5 個(gè)群體中;單倍型Hap 3(8.85%)和單倍型Hap 15(3.98%)的個(gè)體分布于4個(gè)群體中,單倍型Hap 6(8.85%)的個(gè)體分布于3 個(gè)群體中,其余單倍型分布較少。各單倍型在群體中的分布見(jiàn)表1。
5 個(gè)大銀魚(yú)群體總體單倍型多態(tài)性(nh)為(0.833±0.016),核苷酸多樣性(Pi)為(0.00209±0.00020)。每個(gè)群體的單倍型數(shù)(nh)為5~16 個(gè),LHP最少而LHH 最多;單倍型多樣性(Hd)在(0.656±0.051)和(0.939±0.028)之間,核苷酸多樣性(Pi)在(0.00121±0.00010)和(0.00496±0.00150)之間,LHP 多樣性最低而XLC 多樣性最高。Tajima's D 中性檢驗(yàn)值的變化范圍為-1.89133~0.55966,中性檢驗(yàn)結(jié)果顯示只有XLC(P<0.05)顯著不符合中性突變,其他4 個(gè)群體均不顯著,符合中性突變(表2)。
2.3 群體遺傳分化
Cyt b 基因的AMOVA 結(jié)果表明,各群體間的遺傳差異主要來(lái)自群體內(nèi)(93.81%),少數(shù)來(lái)自群體間(6.19%)(表3)。群體間遺傳分化系數(shù)(Fst)為0.06185,各群體間的遺傳分化系數(shù)(Fst)為0.01537~0.13572,除了查干湖與龍虎泡分化不顯著外,其余群體間分化顯著(P<0.05),查干湖和連環(huán)湖、新立城和龍虎泡、二龍山水庫(kù)與龍虎泡和查干湖達(dá)到了極顯著的遺傳分化水平(表4)。
2.4 遺傳距離和系統(tǒng)進(jìn)化樹(shù)的構(gòu)建
利用Cytb 基因序列,采用MEGA 4.0 軟件,根據(jù)Kimura 雙參數(shù)模型計(jì)算各群體之間的遺傳距離(D)。結(jié)果顯示:XLC 和ELS 之間遺傳距離最大(0.0040),LHP 和LHH 之間遺傳距離最小(0.0014)(表4)。根據(jù)各群體之間的遺傳距離構(gòu)建的系統(tǒng)進(jìn)化樹(shù)顯示,CGH 和LHP 聚為一支后與LHH 聚為一支,再與ELS 聚為一支,XLC 獨(dú)立為一支(圖1)。
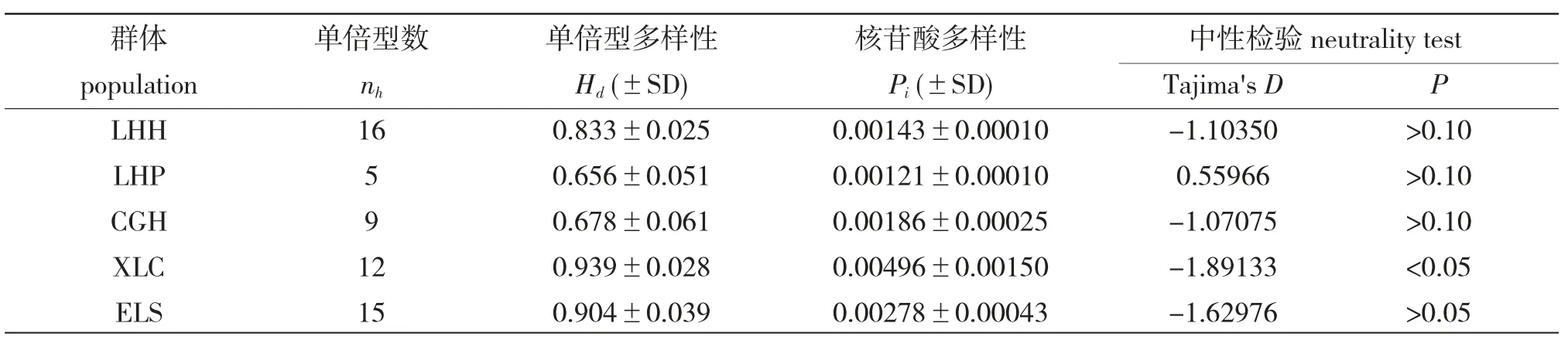
表2 基于線粒體Cyt b 基因的遺傳多樣性Tab.2 Genetic diversity estimates in the five populations of clearhead icefish based on Cyt b gene of mtDNA
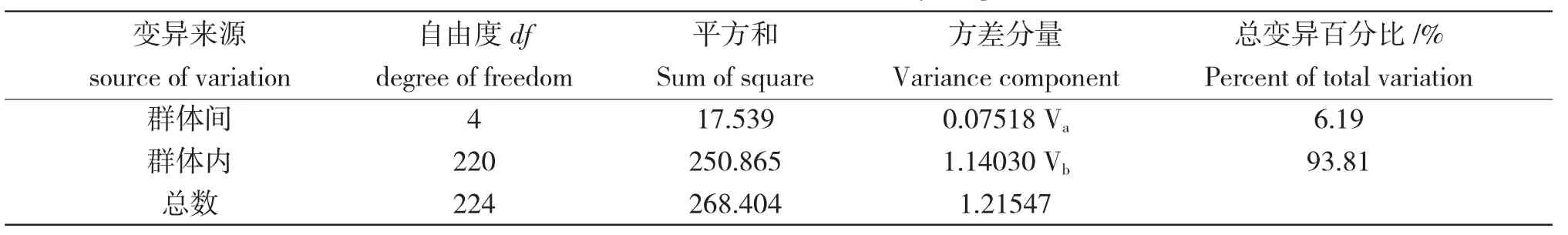
表3 基于線粒體Cyt b 基因的AMOVA 分析結(jié)果Tab.3 The result of AMOVA based on Cyt b gene of mtDNA
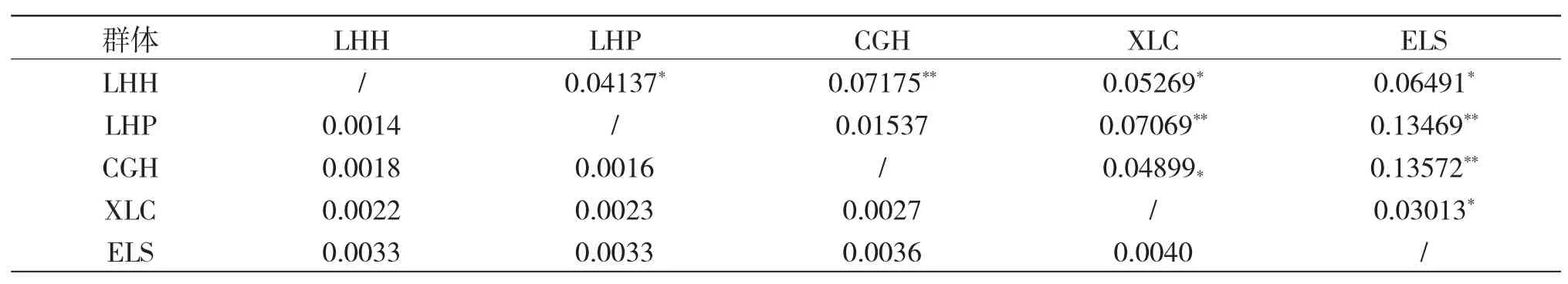
表4 基于Cyt b 基因的各群體間的Fst 和基于Kimura 雙參數(shù)模型計(jì)算的各群體間遺傳距離Tab.4 The Fst among populations based on Cyt b gene and genetic distance calculated based on Kimura 2-parameter model in each population
3 討論
銀魚(yú)營(yíng)養(yǎng)豐富、口感好、經(jīng)濟(jì)價(jià)值高,是我國(guó)重要的出口創(chuàng)匯水產(chǎn)品之一。自20 世紀(jì)90 年代以來(lái),陸續(xù)移植到全國(guó)27 個(gè)省(市)的湖泊、水庫(kù)進(jìn)行養(yǎng)殖[23],其中大銀魚(yú)個(gè)體大、冬季繁殖的生物學(xué)特性,使其具有較強(qiáng)的適應(yīng)能力,特別是對(duì)寒冷有較強(qiáng)的耐受力,而在北方的湖泊水庫(kù)成功移植,在諸多省份形成了產(chǎn)量,獲得了顯著的經(jīng)濟(jì)效益和社會(huì)效益[2]。
線粒體DNA(mtDNA)具有嚴(yán)格的母系遺傳、進(jìn)化速率快、幾乎無(wú)重組等特點(diǎn),被廣泛用于魚(yú)類系統(tǒng)發(fā)育和群體遺傳研究。其中Cyt b 基因作為重要的蛋白質(zhì)編碼基因,進(jìn)化速率適中,能有效反映屬、亞種、種群等不同群體水平的遺傳信息[22],廣泛應(yīng)用于銀魚(yú)系統(tǒng)進(jìn)化及遺傳資源評(píng)估中[9-13]。用Cyt b 基因序列在原產(chǎn)地太湖和洪澤湖大銀魚(yú)群體中分別檢測(cè)到12 個(gè)和7 個(gè)單倍型,單倍型多樣性分別為(0.850±0.045)和(0.775±0.045),核苷酸多樣性分別為(0.00296±0.00017)和(0.00129±0.00010)[11,12]。本研究中,在松嫩平原區(qū)3 個(gè)湖泊和2 個(gè)水庫(kù)大銀魚(yú)群體中共檢測(cè)到38 個(gè)單倍型,單倍型多樣性為(nh)和核苷酸多樣性(Pi)分別為(0.833±0.016)和(0.00209±0.00020),略低于太湖群體而高于洪澤湖群體,其中連環(huán)湖群體和二龍山群體的單倍型數(shù)高于太湖,新立城水庫(kù)和二龍山水庫(kù)的遺傳多樣性指標(biāo)也高于太湖群體,龍虎泡和查干湖群體遺傳多樣性較低。連環(huán)湖是嫩江下游平原型湖泊,本研究的樣本分別采自連環(huán)湖最下游的水體阿木塔泡和牙門喜泡,而所有連環(huán)湖水均經(jīng)阿木塔泡后流入嫩江,因此,上游水源帶來(lái)的大銀魚(yú)增加了該水體大銀魚(yú)的遺傳多樣性。新立城水庫(kù)和二龍山水庫(kù)均為第二松花江支流的水庫(kù),均采用人工投放受精卵的方式維持遺傳多樣性;兩水庫(kù)不僅與松嫩平原湖泊進(jìn)行大銀魚(yú)種質(zhì)交換,也與南方湖泊進(jìn)行了較多的種質(zhì)交換,增加了本地區(qū)其他湖泊不具備的單倍型,因此保持了較高的遺傳多樣性水平[23,24]。龍虎泡和查干湖均為獨(dú)立的水體,與外界水體溝通較少,增加遺傳多樣性的措施也為投放不同來(lái)源受精卵,但本研究顯示二者遺傳多樣性較低,應(yīng)注重引進(jìn)更廣泛水體的大銀魚(yú)受精卵。
AMOVA 結(jié)果表明:大銀魚(yú)遺傳差異主要來(lái)自群體內(nèi)(93.81%),少數(shù)來(lái)自群體間(6.19%)。雖然群體間遺傳分化系數(shù)較低(Fst=0.06185),但是兩兩群體間遺傳分化均達(dá)到顯著水平或極顯著水平,顯示出各水體不同批次移植、投放大銀魚(yú)受精卵的特征;而查干湖與龍虎泡群體分化不顯著,可能是兩水體常態(tài)化交換投放受精卵的結(jié)果。聚類分析結(jié)果表明松嫩平原區(qū)湖泊連環(huán)湖、查干湖、龍虎泡首先聚類,然后再與第二松花江支流的二龍山水庫(kù)和新立城水庫(kù)聚類,體現(xiàn)出嫩江下游平原堿性湖泊遺傳相似度高而松花江兩水庫(kù)與其他各水體相似度均低的特征。
大銀魚(yú)在土著水域正面臨著資源衰退的問(wèn)題,其廣泛移植不僅創(chuàng)造了很好的經(jīng)濟(jì)效益和社會(huì)效益,也增加了我國(guó)這一獨(dú)有物種的遺傳多樣性,這對(duì)增強(qiáng)該物種的可持續(xù)利用具有重要意義。但是,大銀魚(yú)的移植增殖會(huì)明顯抑制其他魚(yú)類種群發(fā)展,特別是小型上層魚(yú)類,如屬Hemiculter sp.魚(yú)類等。因此,認(rèn)為不應(yīng)在水生生物保護(hù)區(qū)水域進(jìn)行大銀魚(yú)移植增殖;同時(shí),在管理大銀魚(yú)種群時(shí)要根據(jù)水體環(huán)境容納量確定大銀魚(yú)卵投放量和親魚(yú)保留量,防止過(guò)度放養(yǎng)和過(guò)度消耗餌料生物資源,保護(hù)其他魚(yú)類資源是大銀魚(yú)種群可持續(xù)發(fā)展和漁業(yè)利用的基礎(chǔ)。
