氮添加對大果木姜子凋落葉分解及養分釋放的影響
2021-01-08 03:37:38劉濟明陳敬忠黃路婷
浙江農林大學學報 2020年6期
陳 夢,劉濟明,陳敬忠,李 佳,黃路婷
(貴州大學 林學院,貴州 貴陽 550025)
氮素在森林生態系統的能量流動與物質循環過程中有著非常重要的作用[1?2]。長期氮沉降以及人類活動干擾等,造成全球氮(N)沉降的增加,對自然生態系統產生了一系列影響[2?3]。研究表明:即使在沒有物理擾動的情況下,長期氮沉降也會通過降低碳周轉率、改變氮的有效性和轉化來影響溫帶森林生態系統的生物地球化學循環[4]。此外,也有研究表明:氮添加會通過改變凋落葉特性(碳、氮、磷變化)對其養分分解速率產生影響[3]。據報道,中國已成為全球第三大氮沉降地區,大氣含氮化合物增長迅速[5]。在未來幾十年中氮沉降繼續增加的背景下,研究氮沉降對森林凋落物養分分解的影響顯得尤為重要[6?7]。凋落物在森林生態系統組成中占據重要地位,它的分解也會對森林生態系統的生物地球化學循環產生相當大的影響[8?9]。凋落物分解不僅是林下土壤有機質形成所需碳源以及土壤養分的主要來源,而且對于土壤質地的構建、肥力的維持、微生物代謝的調控都起重大作用,并且通過這一系列作用進而影響微生物的群落結構[10?11]。大果木姜子Cinnamomummigao又名米槁,系樟科Lauraceae樟屬Cinnamomum常綠喬木,主要分布于中國廣西、云南、貴州三省交界的南北盤江、紅水河流域,為中國特有種。其果實常常被苗族作為治療胃腸道疾病的傳統藥物,并且療效卓越。21世紀初,貴州省科學技術廳以及貴州省民族醫藥研究所確定大果木姜子為貴州省的道地藥材[12]。近年來,隨著其藥用價值的提高,大果木姜子野生資源受到極大破壞,在《中國生物多樣性紅色名錄——高等植物卷》(2013版)已將其列為近危種,野外資源儲量極其有限。目前,相關學者對大果木姜子的研究報道還不是很全面,主要集中在果實精油、脂肪油、揮發油化學成分分析[13]、藥用成分的藥理作用與生物活性[14?15]、栽培技術與病蟲害防治[12]、地理分布[15]、生物生理特性[16?17]等方面。大果木姜子凋落葉的分解對氮沉降增加的響應規律還未見報道。本研究通過野外模擬氮沉降的試驗,探討不同施氮處理對大果木姜子凋落葉養分分解的影響,以期探討大果木姜子凋落葉分解對外源氮添加的響應機制,且為其物質循環機理研究提供理論依據。
1 試驗地概況
貴州省羅甸縣位于云貴高原和丘陵過渡的斜坡地帶,四季呈現春早、夏長、秋遲、冬短的特點,因其特殊的地勢與優良的自然環境被稱為大果木姜子生長的典型生境。試驗樣地設置于羅甸縣逢亭鎮西部的祥林村 6 年生大果木姜子藥材種植基地,地處 25°15′41′N,106°31′03′E,海拔 524 m。屬于亞熱帶季風氣候,年平均氣溫20.6 ℃,≥10 ℃年積溫為6 125.0 ℃,年降水量1 200.0 mm,無霜期335.0~349.5 d,年均日照時數1 448.9 h。試驗區土壤為酸性黃壤,土層厚度≥40 cm。于2017年1月初在試驗地設置12塊大小為3 m×5 m的樣地進行試驗。
2 研究方法
2.1 試驗設計
以2010年中國大氣氮沉降特征分析中貴州的氮沉降總和低于15 g·m?2·a?1為依據,本研究從低到高設置 4 個氮處理,即對照 (ck,0 g·m?2·a?1)、低氮 (N1,5 g·m?2·a?1)、中氮 (N2,15 g·m?2·a?1)和高氮(N3,30 g·m?2·a?1),每處理重復3次。為消除環境異質性的影響,各樣地之間間隔5 m。按照試驗設計要求,每個處理設定樣方面積為3 m×5 m,將各樣方每次所需噴施的NH4NO3溶解于20 L自來水中,采用按壓式噴壺來回均勻噴灑至樣地,ck組噴灑等量的水。于試驗2017年1月進行首次施氮試驗,共噴灑3次,分別于1、5、9月取樣完成,重新放置尼龍袋后進行等量施氮。
2.2 凋落葉的收集與處理
凋落葉分解試驗于2017年1月開始進行,采用尼龍網袋分解方法。準備540個25 cm × 25 cm的分解袋,每個樣地放置45袋。試驗前于羅甸縣大果木姜子林種植基地收集大果木姜子凋落葉帶回實驗室,清洗風干后裝入分解袋中,10.00 g·袋?1。試驗期間放置在12個施氮處理樣地中(分解袋放置前修整空地表面枯枝落葉層,平鋪網袋,使其盡可能接近自然分解狀態。每次放置好尼龍袋后立即噴施氮源)。每隔2個月于各樣地隨機收取3袋分解袋,用刷子清除每次取回分解袋表面的泥垢后,置于75 ℃烘箱烘干至質量恒定,稱其質量用于計算凋落葉質量損失率。然后研磨過孔徑0.5 mm尼龍篩,保存備用。質量損失率(W):W=Xt/X0×100%。其中:X0為凋落葉初始質量,Xt為分解t時間后凋落葉質量。
2.3 凋落葉養分質量分數的測定
凋落葉全氮質量分數采用H2SO4-H2O2靛酚藍比色法測定,全磷質量分數采用鉬銻抗顯色法測定,全鉀質量分數采用HClO4+HNO3消煮-原子吸收分光光度法測定,碳質量分數采用濃硫酸-重鉻酸鉀法測定。凋落葉各元素的殘留率按照下式計算。元素殘留率(R):R=XtCt/X0C0×100%。其中:X0為凋落葉初始質量,Xt為分解t時間后凋落葉的質量,C0為凋落葉初始養分質量分數,Ct為t采樣時間凋落葉養分質量分數。
2.4 數據處理與分析
運用Excel和SPSS 18.0軟件進行初步統計分析,采用最小顯著差法(LSD)多重方差顯著性檢驗凋落葉質量損失率、殘留率和養分質量分數與不同施氮處理間的差異顯著性,利用Olson 負指數衰減模型(y=ae?Kt)對質量殘留率與時間的關系進行擬合,其中:y為質量殘留率(%),a為擬合參數,K為分解系數,t為時間(a)。利用Origin 2018繪制圖表。
3 結果分析
3.1 不同處理對大果木姜子凋落葉質量殘留率的影響
圖1顯示:1?9月各處理凋落葉質量損失率隨著時間的推移表現為迅速遞增趨勢,其中ck凋落葉質量損失率大于其他處理;9月以后各處理凋落葉分解速率趨于平緩。整個分解試驗過程中,N3處理凋落葉質量損失率為0~53.35%,且始終低于其他處理。9月以前,N1、N2處理凋落葉質量損失率顯著高于N3處理(P<0.05),即凋落葉質量損失率從大到小依次為N2、N1、N3;分解結束時(11月),各處理凋落葉質量損失率從大到小依次為 N2、ck、N1、N3。
從表1可以看出:4個處理的擬合模型R2均大于0.900 0,且達到顯著水平(P<0.05),其中N3處理Olson模型的擬合效果最佳。ck處理的凋落葉質量分解系數最大(K=0.085),N3處理最小(K=0.060)。N1、N2、N3處理凋落葉分解95%所需時間分別比ck長0.653、0.312、0.698 a,其中N3處理下凋落葉分解時間所需最長。綜合表明:氮添加延緩了大果木姜子凋落葉的分解。
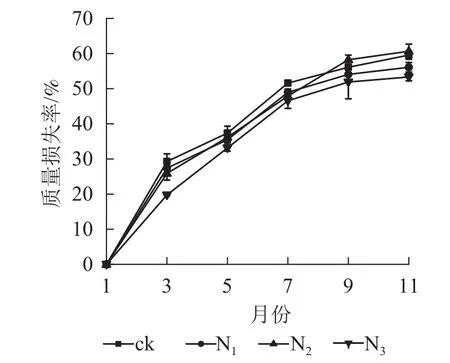
圖 1 不同處理下大果木姜子凋落葉質量損失率Figure 1 Change of mass loss rate and residual rate of leaf litter of C.migao under the condition of different treatments
3.2 不同處理對大果木姜子凋落葉分解過程中碳、氮、磷、鉀的影響
由圖2可知:各處理凋落葉碳質量數總體均呈下降趨勢,且分解前期(1?5月)下降較快,后期(5?11月)下降較慢。各處理凋落葉全氮質量分數變化趨勢基本相同,整體上均呈先增加后降低的趨勢,其中9月最高。各處理凋落葉全磷與全鉀質量分數變化趨勢相似,即在分解初期下降,之后整體上升,最后趨于平穩。
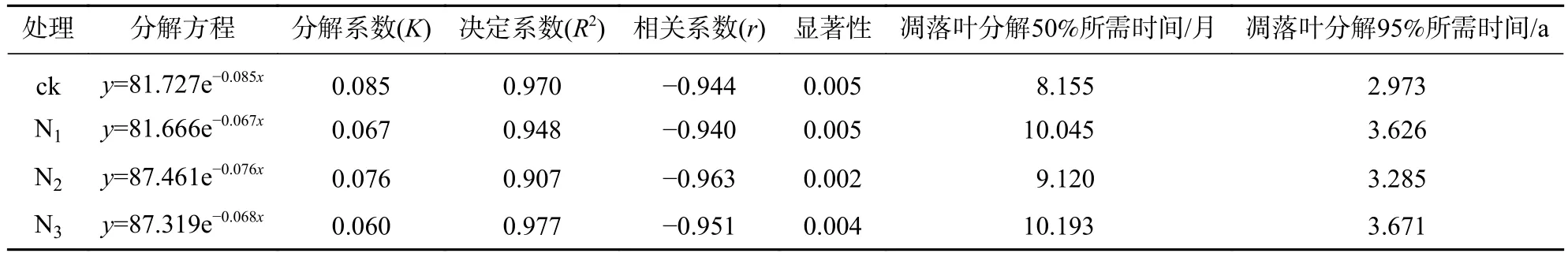
表 1 不同處理大果木姜子凋落葉質量殘留率(y)隨時間(x)變化的Olson模型Table 1 Decomposition rate model of litter residue rate (y) with time (x) under different treatments
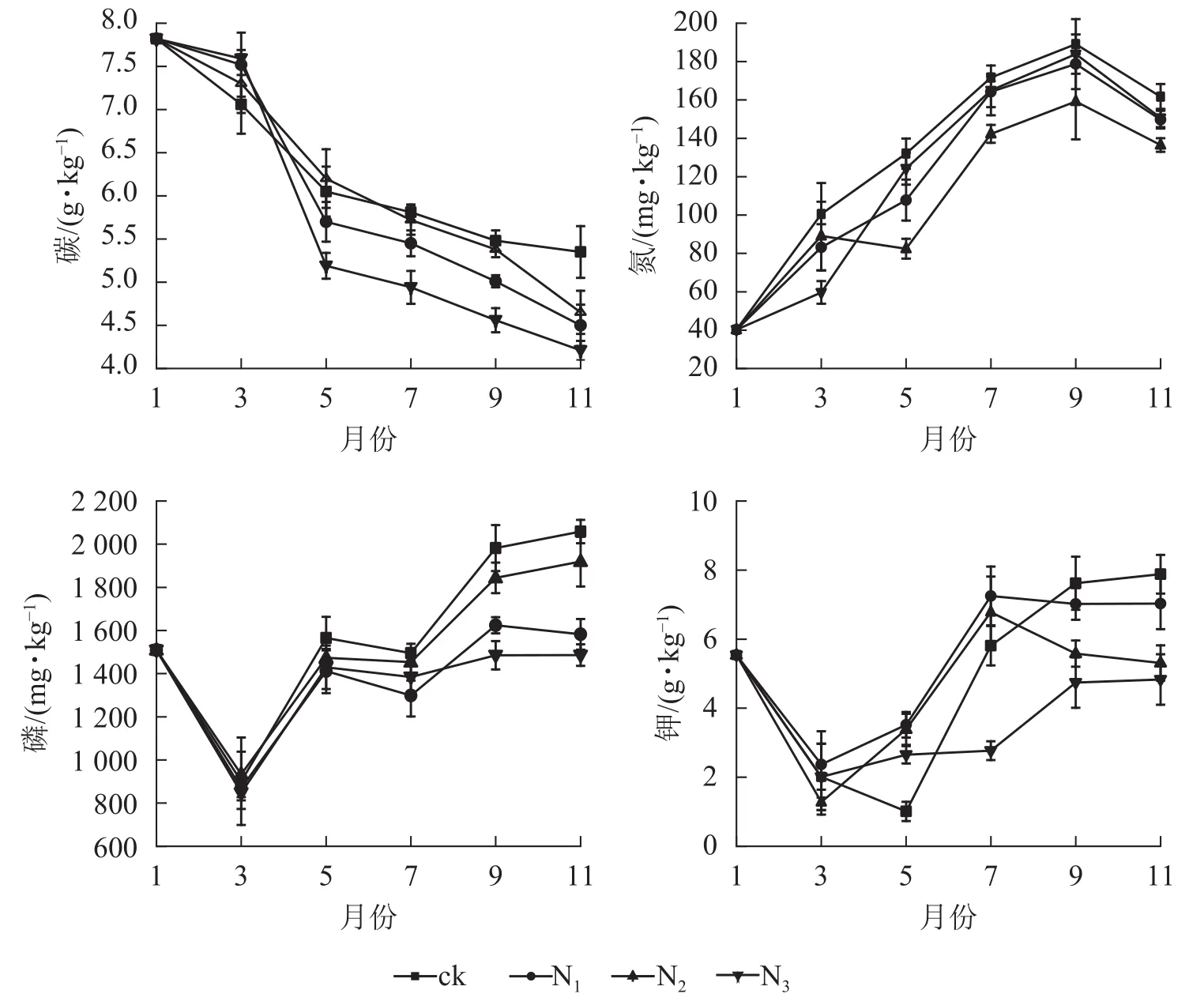
圖 2 不同處理下大果木姜子凋落葉分解過程中養分的變化Figure 2 Change of different treatments on nutrient content of C.migao leaves during decomposition
3.3 不同處理對大果木姜子凋落葉分解過程中元素殘留率的影響
由圖3可以看出:由于碳元素主要以有機形式存在于凋落葉中,受淋溶影響,各處理碳殘留率呈降低趨勢,其中分解各時段3個施氮處理碳殘留率整體上顯著高于ck(P<0.05)。ck的全氮殘留率整體上呈先上升后下降再上升后下降趨勢,各施氮處理的全氮殘留率整體上呈先上升后下降趨勢;1?7月,各處理全磷殘留率呈先下降后升高再下降的趨勢;7月之后,N1、N2、N3處理全磷的殘留率趨于穩定,ck處理全磷殘留率呈先上升后下降趨勢。1?3月,各處理全鉀殘留率迅速降低,3月之后各處理全鉀殘留率總體呈增加趨勢,整體表現為淋溶—富集—釋放模式。
3.4 不同處理對大果木姜子凋落葉分解過程中碳氮比(C/N)的影響
從圖4可以看出:試驗期間各處理C/N為24.60~177.51,整體表現出下降趨勢。分解前期(1?7月)C/N下降速度較快,后期(7?11月)變化總體上比較平穩。在分解各時間段,ck的C/N基本高于其他各施氮處理。在試驗前期(1?7月) ck處理與各施氮處理間有顯著差異(P<0.05);試驗后期(7?11月) ck組與N1、N2處理差異顯著(P<0.05),與N3差異不顯著(P>0.05)。總體來看,添加氮降低了大果木姜子凋落葉的C/N。
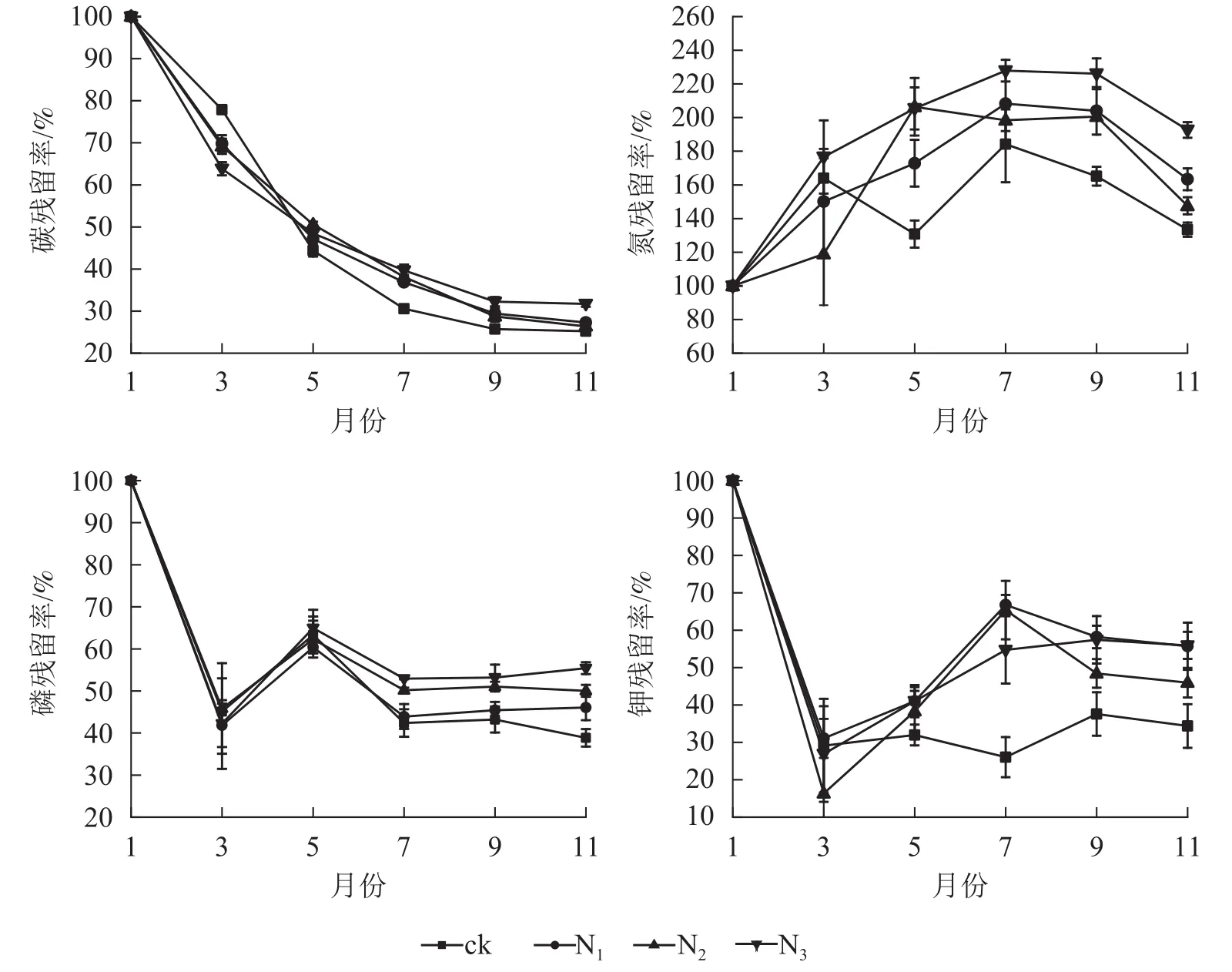
圖 3 不同處理下大果木姜子凋落葉分解過程中元素殘留率的變化Figure 3 Change of different treatments on element residual rate of C.migao leaves during decomposition
4 結論與討論
4.1 不同處理對大果木姜子凋落葉分解質量損失率的影響
凋落葉分解速率的大小受到氣候、凋落葉分解質量、微生物類型及作用等條件的影響[8]。目前,關于氮添加對凋落物分解速率的影響主要分為促進、抑制和無影響3種,這主要取決于施氮量的高低及施氮時間的長短。O’CO NNELL[18]分析了施加外源氮對桉樹Eucalyptus凋落物分解及養分含量變化的影響。研究結果表明:施氮處理下凋落物分解釋放速率變慢。鄭欣穎等[19]模擬外源氮添加對火力楠Micheliamacclurei凋落葉分解影響,結果顯示:氮添加會促進凋落葉的分解。樊后保等[20]通過對杉木Cunninghamialanceolata人工林凋落物施加外源氮,結果出現了“低促高抑”現象。文海燕等[21]對長芒草Stipabungeana和阿爾泰狗娃花Heteropappusaltaicus進行凋落葉模擬氮沉降試驗。結果發現:氮添加對2種植物凋落葉分解無影響。
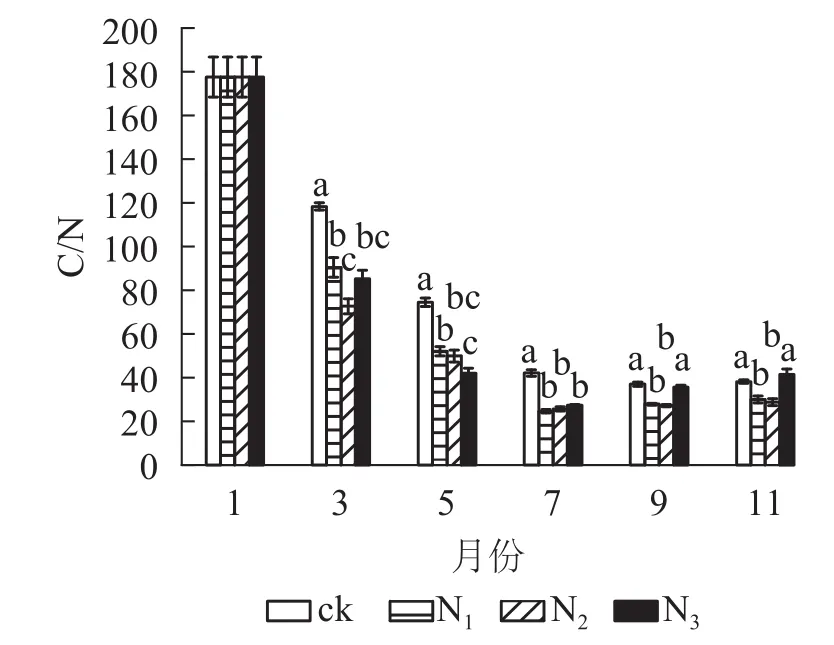
圖 4 不同處理下大果木姜子凋落葉分解過程中C/N的變化Figure 4 Change of different treatments on C/N during decomposition of C.migao leaves
本研究結果顯示:氮添加對大果木姜子凋落葉分解具有一定的抑制作用。大果木姜子凋落葉質量年損失率表現為ck高于各施氮處理,其中N3處理質量損失率最低。這是因為試驗過程中施加外源氮的量超過了試驗樣地土壤所需氮的閾值,從而使凋落葉分解過程中起主要作用的微生物分解效率降低,進而對凋落葉的分解表現出抑制作用[22?23]。凋落葉分解本身具有的養分特性也會使凋落葉在不同分解階段表現出不同規律,通常為前期分解較快,后期變慢。這是因為分解初期凋落物自身養分含量較高,可通過養分流失調控分解速率;隨著養分的流失,后期則通過難分解的木質素和纖維來調控養分分解,所以分解速率表現為前期大于后期,且隨著時間推移逐漸變慢[21?23]。在本研究前期(3?9月),大果木姜子凋落葉質量損失率迅速增加,主要是因為此時凋落葉的質量大,易分解物質含量高,導致凋落葉分解速率隨著易分解物質的分解而加快;試驗后期(9?11月)分解速率緩慢且質量損失率趨于平緩,這與后期凋落葉中易分解物質減少,難分解物質含量升高有關[24?25]。
4.2 不同處理對大果木姜子凋落葉分解過程中養分質量分數的影響
土壤是植被生長過程中主要礦質營養的來源,而凋落物分解釋放的養分歸還是土壤肥力的重要來源(植物生長發育過程中從土壤中吸收的養分70%~90%得益于凋落物的分解釋放)[26],因此,在森林生態系統中,凋落物的分解在植被養分循環過程中起著至關重要的作用。碳元素作為地球一切生命的基本單元,既是植物生長發育的基礎又是維持植物生活的能量來源,氮和磷元素是氨基酸、核苷酸的重要組成元素,鉀元素是調節細胞內外適宜滲透壓和體液酸堿平衡,參與胞內糖和蛋白質代謝的重要陽離子。4種元素都是植物生命過程中不可缺少的[27?28]。研究表明:植物生長過程所需的養分分別有70%~80%的氮、65%~80%的磷和30%~40%的鉀來自凋落物的分解釋放[29]。
本研究中,5?11月各施氮處理碳質量分數顯著低于ck(P<0.05),而氮質量分數表現相反。這是因為外源氮的添加使凋落葉中碳氮轉化酶活性發生改變而影響兩者的質量分數,且氮添加抑制了纖維素等物質的降解從而使凋落葉分解過程出現“碳封存”狀態。凋落葉分解過程中,外界氮源的輸入以及原生質素伴隨自身固氮菌的侵入逐漸增加使氮質量分數增加[20],因此,C/N隨著分解時間變長而總體下降。試驗期間,凋落葉中全磷、全鉀變化不穩定,各處理差異顯著。11月凋落葉全磷質量分數表現為N1、N2、N3處理高于ck,且N2、N3差異顯著(P<0.05),說明氮添加促進了大果木姜子凋落葉中全磷質量分數的積累,且施加氮的量越大表現出的差異越顯著(P<0.05)。這與國內外多數研究結果相同,如BERG等[30]模擬氮沉降對樟子松Pinussylvestris凋落葉養分分解影響研究表明:氮添加促進了凋落葉中全氮、全磷質量分數的積累。肖銀龍等[31]模擬氮沉降對苦竹Pleioblastusamarus林凋落葉養分分解的研究同樣表明:氮添加促進全氮、全磷的積累。本研究中,11月N1和N2處鉀質量分數高于ck,而N3處理低于ck,說明添加過多的氮對凋落葉中全鉀具有抑制作用,這是因為氮添加到土壤后,大量的氮以NO3?的形式淋失,為了達到土壤中養分電荷平衡,必然會帶走等量的鹽基離子(K+)[32?33]。
凋落物中的C/N一般比較高,不利于微生物的活動,而增施氮肥后有利于微生物數量的增加,從而促進凋落物的分解。本研究中土壤本身氮質量分數較高,再增施氮肥使其超過了“臨界值”,打破了土壤中養分平衡,從而氮添加對大果木姜子凋落葉分解表現出抑制作用[22]。
綜上所述,在氮添加的情況下,不利于大果木姜子凋落葉分解及養分的釋放,其中N3處理作用更顯著。凋落物分解后養分歸還于土壤,再次作用于植物,但土壤養分輸入對氮添加作何響應,還需進一步進行模擬探究。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
當代陜西(2021年2期)2021-03-29 07:41:24
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
汽車觀察(2016年3期)2016-02-28 13:16:26
