二十四元大環內酯生物合成研究進展*
2021-01-14 08:44:48甘雨滿黃炳耀唐振洲高程海
廣西科學 2020年5期
關鍵詞:生物
甘雨滿,黃炳耀,唐振洲,林 霄,高程海
(廣西中醫藥大學海洋藥物研究院,廣西南寧 530200)
0 引言
傳統抗生素的無節制使用,導致病原菌抗藥性問題日益突出,挖掘新型結構、高活性的抗生素已成為解決病原菌抗藥性問題的重要手段。隨著從陸地資源挖掘新型化合物的潛力下降,人們逐漸將目光轉向海洋生物資源,種群豐富的海洋微生物將是挖掘新型化合物的聚寶盆。海洋生境是一個復雜的生態系統,其高鹽、無光、低溫(局部高溫)、低營養及高壓的特殊環境孕育了海洋生物物種的多樣性。物種種類優勢以及復雜的海洋環境造就了微生物獨特的代謝方式,賦予了人類從海洋微生物獲取新型化合物的可能。截至2017年,已完成分離鑒定的海洋微生物來源的新型天然產物達1 490種,來源微生物集中在細菌、霉菌、放線菌和藍藻[1]。二十四元大環內酯化合物Macrolactins (MLNs)主要來源于海生Bacillussp.和Actinomadurasp.,包含超過32種Macrolactin (MLN)單體化合物[2]。自Fenical首次從深海芽孢桿菌中分離鑒定MLN A-F 6種單體以來,MLNs因具備抗病毒、抗腫瘤和抗病菌等多種生物抑制活性而備受關注。MLN A 除了可抑制HIV外,還對哺乳動物皰疹病毒和腫瘤細胞表現出抑制作用;MLN F 對鯊烯合成酶具有抑制作用,可作為調控膽固醇水平的潛在藥物[3,4]。在抑菌活性方面,MLN W可有效抑制革蘭氏陽性菌和部分革蘭氏陰性菌[5]。MLN T不僅能夠對常見病原菌耐甲氧西林金黃色葡萄球菌(Methicillin-resistantStaphylococcusaureus)表現出抑制作用,而且對植物致病菌交鏈孢(Alternariaspp.)和稻瘟霉(Pyriculariaoryzae)也表現出較明顯的抑制效果[6]。可見MLNs是一類具有廣譜生物抑制活性的化合物,在治療疾病、防治農業病蟲害生物等領域具有廣闊的應用潛力[7]。但野生菌株代謝產生的MLNs產量較低,限制了該類化合物的進一步研究。 鑒于此,本文對MLNs生物合成研究現狀進行綜述,包括MLNs合成基因簇、代謝菌株遺傳改造、發酵工藝優化等,以期為MLNs的批量生產提供研究資料。
1 MLNs合成基因簇
MLNs是一類聚酮化合物,是由生物體通過聚酮合酶(Polyketide Synthase,PKS)基因簇逐級縮合產生的天然化合物。根據結構和功能的差異,PKS酶系可分為Ⅰ型、Ⅱ型和Ⅲ型。Ⅰ型PKS是由多個模塊按線性排列而成的酶催化體系,每個模塊含有一個特有的催化結構域,常用來催化形成具有大環結構的硫酯化合物[8];Ⅱ型PKS是一個多功能酶復合體,由一套獨立的4—6個可重復使用的酶組成,每一種酶在重復的反應步驟中可多次催化相同的反應[9];Ⅲ型PKS與前兩種類型的明顯區別是,可在不需要ACP的情況下直接催化泛酞輔酶A的縮合[10]。
隨著生物信息學的快速發展,生物遺傳密碼被不斷破譯[11]。Borriss 研究團隊通過分析BacillusamyloliquefaciensFZB42基因組,首次確認了Ⅰ型PKS基因簇參與MLNs合成[12]。整個基因簇大小約54 kb,共由11個重復模塊組成,每個模塊至少包括酰基轉移酶、酮基合成酶和酰基載體蛋白3個元件。每個元件在 MLNs碳骨架的合成過程中承擔著不同的生物功能:酰基轉移酶負責底物的激活和運轉,酮基合成酶負責延伸聚酮鏈,酰基載體蛋白負責將完成延伸2個碳原子的聚酮鏈運轉到下一模塊。MLNs聚酮鏈的合成過程與脂肪酸碳骨架的合成過程類似,具體過程如下:延伸單元丙二酰輔酶A被酰基轉移酶激活后,在酮基合成酶作用下,激活態的丙二酰輔酶A與酰基轉移酶結合的底物分子發生克萊森(Claisen)反應,底物延伸2個碳原子;延伸產物由酰基載體蛋白運轉到下一模塊的酰基載體蛋白,完成一次二碳單元的延伸。經過11輪重復“激活、延伸、運轉”3個步驟,完成二十四元環骨架的合成(圖1)。隨后,脫水酶、酮還原酶、烯醇還原酶進一步對二十四元環骨架側鏈基團進行修飾,修飾模塊多樣化決定了MLNs化合物結構的多樣化[13]。
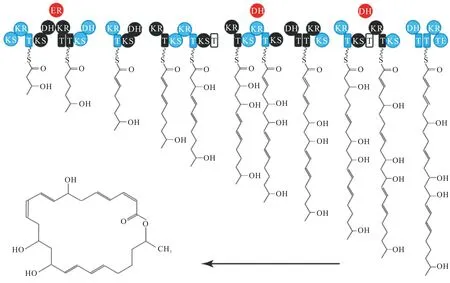
圖1 Macrolactin A生物合成基因簇[12]
2 PKS基因簇多樣化
PKS基因簇根據結構和功能可分為Ⅰ型、Ⅱ型和Ⅲ型,各類型的PKS基因簇由于序列、調控元件等因素差異也呈現多樣化狀態。海洋生態系統中微生物種類豐富,孕育著多樣化的基因資源,董曉毅[14]根據Ⅰ型PKS保守序列設計簡并引物,擴增海水、沿岸土壤、海洋沉積物等不同生境的微生物基因組,獲得多樣化的Ⅰ型PKS,為挖掘新型結構的MLNs化合物提供理論基礎。在PKS結構分析過程中,研究者發現酮縮合酶(KS)片段可能來源于PKS-NRPS雜合基因簇,Moffitt等[15]研究表明,PKS-NRPS雜合基因簇的KS結構域在活性半胱氨酸N-末端方向的第三個氨基酸殘基被谷氨酰胺替代,突變后的KS可以以氨基酸作為起始單元參與生物合成。據文獻報道,PKS-NRPS雜合基因簇包括兩種類型,一是整體PKS模塊和整體NRPS模塊相連接(介導雜合化合物的合成),二是由PKS和NRPS的單一模塊交替相接形成雜合體[14]。由于PKS-NRPS雜合基因簇的特殊性,對PKS-NRPS雜合基因簇的深入研究將有利于新型MLNs化合物的挖掘[16]。
此外,陸生和海生微生物PKS基因簇也表現出差異,宋婧文[17]對陸生和海生MLNs代謝菌株的基因組進行比較分析,發現海生微生物PKS基因簇啟動子區缺失25個堿基,根據MLNs代謝水平與代謝時間的線性關系,研究者推斷這段缺失序列可能參與MLNs合成調控。
3 人工設計PKS基因簇
PKS各個模塊按線性排列,每個模塊均具有特定的生物學功能,PKS模塊重排有利于合成“人工設計”的天然產物。在酮基合成酶作用下,激活態的丙二酰輔酶A與酰基轉移酶結合的底物分子發生克萊森反應,底物延伸2個碳原子,通過多次的克萊森反應完成目標化合物的碳骨架延伸,最后在硫酯酶的作用下長鏈碳骨架閉環合成環形結構。可見,PKS模塊的排列順序決定了化合物骨架的碳原子數目和結構,同理,改變這些模塊的排列順序也可導致新化合物的生成[18]。PKS模塊這種一一的對應關系,為編輯PKS基因簇基因(具體操作包括結構域或模塊的互換、突變、插入、刪除)提供了可能。但是對PKS的模塊編輯面臨不少問題[18-20],一是對野生菌株而言導入目的DNA片段具有一定挑戰性,尤其是部分不可培養或者生長極其緩慢的菌株;二是PKS基因簇具有不易操作性,具體表現為基因簇往往太長(35-200 kb)、沒有保守位點,以及模塊重復不利于定點編輯。針對PKS基因簇不易操作性難題,Menzella等[18]在模塊和結構域兩端插入限制性酶切位點,模塊間或結構域間通過限制性酶切位點重新組合,高效獲得重構PKS基因簇(圖2),接近一半的重構基因簇表現出正常的基因簇生物學功能,并獲得結構多樣的化合物。

圖2 PKS模塊組裝[18]
限制性酶切位點策略很好地解決了PKS基因簇重構的難題,但在天然宿主菌種中表達PKS基因簇,產量往往較低,不利于檢測和后續的生物學研究。異源表達是快速、高效表達目的基因的重要手段,常用的異源表達宿主菌包括枯草芽孢桿菌、大腸桿菌、畢赤酵母等,這些異源宿主菌具有易于培養、遺傳背景清晰、遺傳操作技術成熟等優勢,便于構建異源表達工程菌株。除此之外,PKS基因簇異源表達載體構建需要考慮的因素包括選擇具備相當容量的載體、密碼子優化等問題。目前已成功表達PKS基因簇的異源宿主菌包括酵母菌、大腸桿菌、天藍色鏈霉菌、變鉛青鏈霉菌、弗氏鏈霉菌、糖多孢紅霉菌等[14]。
4 從遺傳水平調控MLNs生物合成
從遺傳水平對微生物的MLNs合成途徑進行調控,一方面可以挖掘新型結構的MLNs化合物,另一方面也是提高已知MLNs化合物代謝水平的主要手段。高效的遺傳轉化手段是進行遺傳操作的重要基礎,但和實驗室菌株相比較,從環境中篩選獲得的野生微生物由于細胞壁構造特殊且遺傳背景不清晰,適用于實驗室菌株的遺傳操作技術對于其往往效率低下。針對野生菌株重新建立一套高效的遺傳操作技術對于微生物遺傳育種而言是必要的。Liu等[21]對一株產MLNs的海洋芽孢桿菌遺傳轉化參數進行優化,通過評價轉化體系、培養基、目的DNA去甲基化、滲透壓保護劑以及其他化學成分對海洋芽孢桿菌遺傳轉化效率的影響,首次建立了海洋芽孢桿菌的高效轉化體系,轉化效率可達7.1×104cfu/μg質粒,為MLNs代謝菌株遺傳改造奠定了重要基礎。
Borriss研究組于2007年首次闡明了MLNs合成基因簇[12],但一直以來其分子調控機理并不清晰,合成關鍵基因也鮮有報道,阻礙了從遺傳水平定向調控MLNs生物合成的研究進程。MLNs合成模塊中,反式酰基轉移酶負責延伸單元丙二酰輔酶A的運轉,理論上提高丙二酰輔酶A的運轉速率有利于提高MLNs合成水平。劉揚等[22]過度表達反式酰基轉移酶BmmA,結果顯示MLNs A的產量提高了約0.6倍,說明丙二酰輔酶A基因在MLNs生物合成過程中參與調控MLNs代謝。改變PKS基因簇主要元件的表達水平可調控MLNs代謝水平,改變修飾元件的表達水平也能影響MLNs的代謝。BmmGT1基因負責海洋芽孢桿菌B-9987的MLNs代謝產物的糖基化修飾,BmmGT1基因過度表達后,非糖基化MLNs水平迅速降低,并獲得5種糖基化的MLNs衍生物[23]。
MLNs生物合成過程的調控機制和主要關鍵基因尚不清晰,不利于應用現代分子生物學技術構建優良工程菌株。傳統微生物育種手段,如誘變[24]、適應性馴化[25,26]、Genome shuffling[27]等,其突出優點是不需要對目標菌株遺傳背景有清晰的了解。Yi等[28]運用ARTP誘變育種技術對海洋芽孢桿菌進行誘變,基于菌株的生長系數篩選獲得優良突變菌,其MLNs代謝水平提高52.8%。研究者進一步對突變菌株轉錄組進行研究,156個差異表達基因主要涉及5個生物過程,包括組氨酸代謝、核黃素代謝、非核糖體肽結構、脂肪酸降解和脂肪酸代謝。這些基因組學研究數據為探索MLNs代謝調控機制提供了理論支撐,其中特別值得關注的生物途徑是脂肪酸代謝,因為細胞膜脂肪酸在提高細胞抗生素抗性方面發揮重要作用[29,30]。MLNs是一類廣譜抗生素,當其濃度累積到一定程度后將不可避免地影響代謝菌株的生理狀態,而提高細胞膜脂肪酸濃度可能有利于高濃度的MLNs積累。
5 發酵優化提高MLNs代謝水平
野生菌株的MLNs代謝水平低,成為制約其進行藥物開發研究的主要瓶頸。微生物培養法獲得大量目標化合物需要滿足兩個基本條件:獲得代謝性能優良的菌株和擴大培養。擴大培養的效果常受培養基組分、溶氧、pH值、溫度等因素的影響,因此基于實驗結果和經濟因素考慮,在開展擴大培養前需要對培養條件進行優化。響應面法是發酵優化的常用技術手段,在提高MLNs產量方面也取得比較理想的結果[31-34],目前已報道的MLNs發酵優化最高產量為851 mg/L[35]。
微生物只有進入生長穩定期才開始大量合成聚酮化合物,然而進入穩定期后,細胞對初級代謝產物的需求顯著下降[36],此時作為初級代謝產物的中間代謝產物丙二酰輔酶A是否與MLNs的合成存在直接聯系,尚未明確。王明璐[34]嘗試在培養基中添加乙酸鈉、丙酸鉀營養成分,通過生物轉化以維持胞內丙二酰輔酶A濃度,但MLNs產量并未受到影響,而添加低濃度的豆油則能明顯促進MLNs代謝。Wang等[36]結合基因組學研究闡明豆油和聚酮化合物合成間的直接聯系,發現菌株在對數期細胞生長的同時還積累三酰甘油(豆油主成分),進入穩定期后胞內三酰甘油降解機制被激活,為穩定期細胞的聚酮化合物合成提供前體化合物以及還原力。
6 展望
MLNs具有廣譜生物抑制活性,在醫藥和農業具有潛在的應用前景。目前針對改善菌株MLNs代謝水平的相關問題已開展了部分工作,但受限于MLNs代謝調控機制、關鍵基因不清晰,菌株MLNs代謝水平低的局面至今仍然沒有得到改善,一定程度上限制了MLNs的生物活性研究以及市場推廣。Ⅰ型PKS基因簇由多個模塊組成,這些基因共同受一個操縱子調控表達。因此,對操縱子的關鍵元件(如啟動子)進行改造,通過改造一個位點實現對整個PKS基因簇的表達進行定向調控,這方面的研究有望進一步改善MLNs代謝水平,也能避免PKS基因簇基因元件因重復性而不利于遺傳操作、可能涉及的多位點操作等難題。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54