廣西5種常見星蟲動物的線粒體基因組特征和系統(tǒng)進化分析*
2021-01-14 08:45:04鐘聲平劉永宏黃亮華黃國強陳秀荔
廣西科學 2020年5期
鐘聲平,蔣 艷,劉永宏,黃亮華,黃國強,陳秀荔
(1.廣西海洋研究所有限責任公司,廣西海水養(yǎng)殖新品種繁育工程技術(shù)研究中心,廣西南寧 536000;2.廣西中醫(yī)藥大學海洋藥物研究院,廣西南寧 530200;3.廣西水產(chǎn)科學研究院,廣西南寧 530021)
0 引言
星蟲動物(Sipuncula)是一類圓筒狀不分節(jié)、具體腔、營穴居生活的海洋蠕蟲,是海洋動物獨有的門類。目前,已有研究報道的星蟲動物超過238種,它們廣泛分布于世界各大洋中,從熱帶海域到極地海域,從深海區(qū)到潮間帶[1]。中國沿海已發(fā)現(xiàn)的星蟲動物有2綱4目6科41種,從南至北均有分布,南方熱帶海域的種數(shù)最多[2,3]。其中,廣西北部灣海域常見星蟲動物為裸體方格星蟲(Sipunculusnudus)、可口革囊星蟲(Phascolosomaesculenta)和厥目革囊星蟲(P.scolops)等,它們生物資源總量大,肉質(zhì)鮮美,營養(yǎng)和藥用保健價值高,是當?shù)貜V受歡迎的海產(chǎn)珍品[4,5]。然而,隨著市場價格的逐年升高,廣西北部灣星蟲動物遭到過度采捕,再加上棲息地環(huán)境被破壞,其野生資源急劇減少[6,7]。星蟲動物的進化地位至今仍存在爭議,其物種形態(tài)鑒別特征容易受到環(huán)境因素的影響,再加上相似物種的趨同進化,往往導致星蟲物種分類的錯誤和混亂[8]。因此,急需開展星蟲動物遺傳多樣性、生物條形碼和系統(tǒng)進化分析等種質(zhì)資源保護和可持續(xù)開發(fā)利用方面的基礎(chǔ)研究。
線粒體基因組作為核外遺傳物質(zhì),具有拷貝數(shù)量多、母系遺傳和進化速率快等特點[9],隨著高通量測序技術(shù)的快速發(fā)展,越來越多的研究開始應(yīng)用線粒體基因組開展后生動物分子系統(tǒng)進化和種群遺傳多樣性研究[10,11]。相比單個線粒體基因,線粒體基因組具有更豐富的遺傳進化信息,其基因排列重組也蘊含豐富的進化信息,在解決后生動物系統(tǒng)進化關(guān)系和種群遺傳多樣性的研究中具有較高的應(yīng)用價值[12]。然而,針對星蟲動物門線粒體基因組的研究長期滯后,2009年才有首個線粒體基因組見報道[13],至今Genbank上僅有3種星蟲動物線粒體基因組序列可供公開下載[14],遠不能滿足星蟲動物系統(tǒng)進化和遺傳多樣性的研究需求。為探明廣西北部灣常見星蟲動物線粒體基因組信息,本研究應(yīng)用高通量測序技術(shù)測定5種星蟲動物線粒體基因組序列,并統(tǒng)計分析其遺傳變異信息和系統(tǒng)進化關(guān)系,為廣西北部灣星蟲動物漁業(yè)資源多樣性調(diào)查和保護提供分子遺傳數(shù)據(jù),并為星蟲動物系統(tǒng)進化研究提供科學參考。
1 材料與方法
1.1 樣品采集和鑒定
研究樣品為裸體方格星蟲、邱米管體星蟲(Siphonosomacumanense)、可口革囊星蟲、厥目革囊星蟲、類革囊星蟲(Phascolosomasimilis)等5種廣西北部灣常見星蟲動物,采自廣西北海市銀海區(qū)和鐵山港區(qū)近海潮間帶灘涂。每個物種采集6個以上個體帶回實驗室,解剖后參考文獻[15]開展形態(tài)鑒定,確定種類后采集體壁肌肉組織放置于-80℃超低溫冰箱保存。
1.2 基因組DNA提取和高通量測序
參考陳建琴等[16]的方法略做修改,每個個體取肌肉組織約30 mg,采用TaKaRa MiniBEST Universal Genomic DNA Extraction Kit (TAKARA BIO INC.)技術(shù)從肌肉組織提取基因組DNA,并采用瓊脂糖凝膠電泳法檢測基因組DNA降解程度與蛋白質(zhì)污染程度,同時使用NanoDrop 2000超微量分光光度計檢測基因組DNA濃度和質(zhì)量。選取符合高通量測序要求的DNA樣品,同一物種的6個個體基因組DNA混合后,用干冰保存送往深圳華大基因股份有限公司開展簡化基因組建庫測序。測序文庫片段長度為350 bp,使用Illumina PE150雙末端測序平臺完成簡化基因組測序。
1.3 線粒體基因組序列組裝和基因注釋
采用Trimmomatic軟件對原始測序數(shù)據(jù)進行質(zhì)量篩選[17],以GenBank上已經(jīng)公布的法國群體的裸體方格星蟲線粒體基因組序列(登錄號:NC_011826)作為參考序列,使用MITObim軟件完成廣西北部灣5種常見星蟲動物線粒體基因組序列組裝[18]。利用在線的線粒體基因注釋工具tRNAscan-SE、MITOS等分析預測線粒體蛋白質(zhì)編碼基因(PCGs)序列[19]、核糖體RNA基因序列(rRNAs)、轉(zhuǎn)運RNA基因序列(tRNAs)的編碼區(qū)域和結(jié)構(gòu)特征,完成線粒體基因組序列的識別和注釋。
1.4 線粒體基因組遺傳變異信息和系統(tǒng)進化分析
采用Clustal X軟件對5種廣西北部灣常見星蟲動物線粒體基因組蛋白質(zhì)編碼基因進行多重序列比對,使用DnaSP v6和PAML等軟件統(tǒng)計分析線粒體基因組蛋白質(zhì)編碼基因變異位點數(shù)、非同義替換率(Ka)和同義替換率(Ks)等遺傳變異信息。從GenBnak下載30個物種的線粒體基因組序列,包括5個星蟲動物、7個環(huán)節(jié)動物、2個腕足動物、2個線蟲動物、6個軟體動物、1個輪形動物、3個刺胞動物、2個螠蟲動物、2個扁盤動物,再加上本次獲得的5個星蟲動物的線粒體基因組,使用PhyML 3.0軟件的最大似然法(ML)構(gòu)建系統(tǒng)發(fā)育樹的最佳進化模型[20],分析星蟲動物的系統(tǒng)進化。
2 結(jié)果與分析
2.1 線粒體基因組基因序列特征
星蟲動物肌肉組織基因組DNA提取結(jié)果顯示:樣品輕微降解,DNA濃度高于100 ng/μL,符合建庫測序要求。高通量測序共產(chǎn)生原始數(shù)據(jù)42 G,其中堿基測序質(zhì)量高于Q20的數(shù)據(jù)超過90%,經(jīng)過質(zhì)量篩選去除低質(zhì)量測序堿基,使用MITObim軟件組裝拼接5個星蟲動物的線粒體全基因序列。
基因組裝和基因注釋結(jié)果顯示,廣西北部灣的裸體方格星蟲、邱米管體星蟲、可口革囊星蟲、厥目革囊星蟲和類革囊星蟲的線粒體基因組序列全長分別為15 375,15 917,15 494,15 515,16 237 bp,線粒體基因組堿基組成均呈AT偏好,其A+T含量分別為57.5%、60.8%、65.6%、65.4%和63.5%。可口革囊星蟲、厥目革囊星蟲、類革囊星蟲的A+T含量高于裸體方格星蟲和邱米管體星蟲。注釋后的線粒體基因組序列上傳至GenBnak,其接收號分別為MG873457、MN813483、MG873458、MT239480和MN813482。方格星蟲屬裸體方格星蟲線粒體基因組由37個基因組成(圖1),包括13個蛋白質(zhì)編碼基因、22個tRNAs和2個rRNAs,與無脊椎動物線粒體基因組原始基因組成一致。然而,管體星蟲屬的邱米管體星蟲(圖2)與可口革囊星蟲屬的可口革囊星蟲(圖3)、厥目革囊星蟲(圖4)、類革囊星蟲(圖5)這4種星蟲的線粒體基因組由38個基因組成,比裸體方格星蟲線粒體基因組多一個重復的tRNAMet。此外,本研究中廣西北部灣5種常見星蟲的線粒體基因組的基因均編碼在H鏈上,星蟲動物的基因排列具有高度的保守性,存在3個高度保守的基因排列區(qū)塊:(1)nad6-cob-tRNAPro;(2)nad4l-nad4-tRNALeu;(3)nad1-tRNATrp-atp6-tRNAArg-tRNAHis-nad5。蛋白質(zhì)編碼基因和2個rRNAs基因排列順序保持一致,僅有部分tRNAs發(fā)生易位。
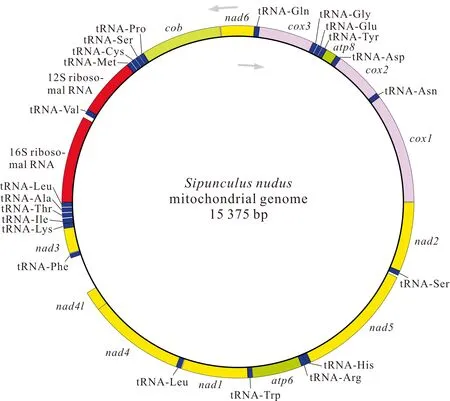
圖1 裸體方格星蟲線粒體基因組的基因圖譜
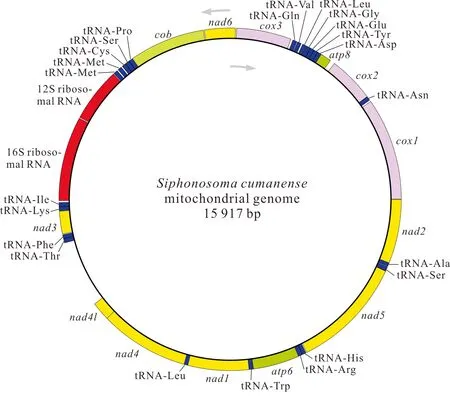
圖2 邱米管體星蟲線粒體基因組的基因圖譜
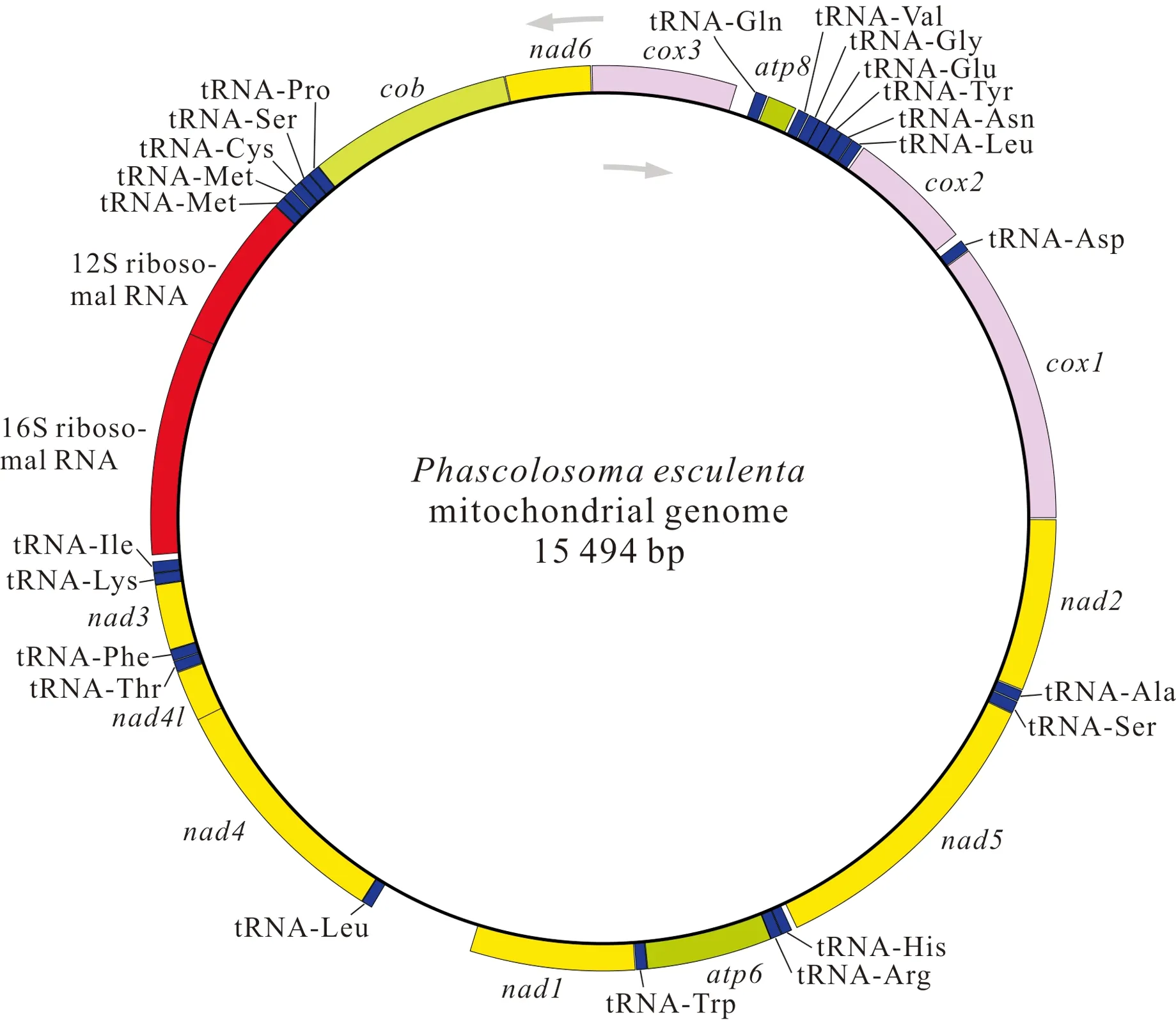
圖3 可口革囊星蟲線粒體基因組的基因圖譜
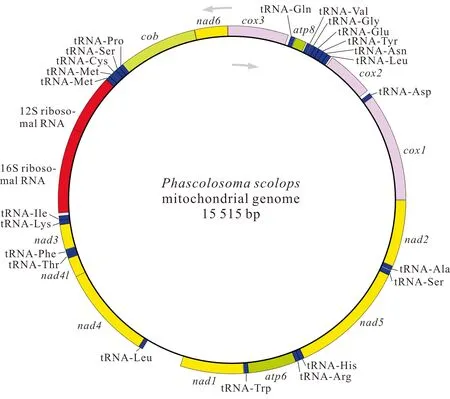
圖4 厥目革囊星蟲線粒體基因組的基因圖譜
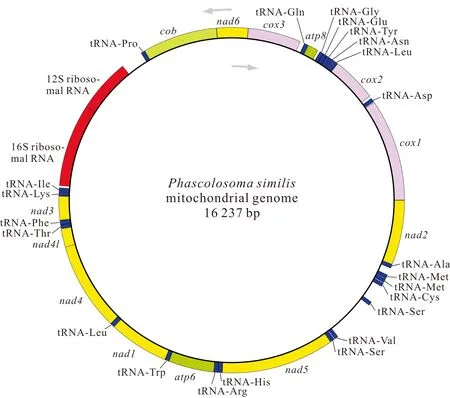
圖5 類革囊星蟲線粒體基因組的基因圖譜
2.2 線粒體基因組遺傳變異信息
采用DnaSP v6軟件,統(tǒng)計分析5種星蟲動物線粒體15個主編碼基因(13個蛋白質(zhì)編碼基因和2個rRNAs基因)的變異位點,結(jié)果顯示,cox1基因最保守,遺傳變異水平最低,為40.53%;其次是cox2和cob基因,遺傳變異水平分別為46.57%和46.83%;nad6、nad4、nad5和nad2等4個線粒體NADH脫氫酶亞基基因遺傳變異水平較高,為60%—70%;atp8基因遺傳變異水平最高,為75.16%(表1)。
采用PAML軟件,分析蛋白質(zhì)編碼基因非同義替換率(Ka)和同義替換率(Ks),結(jié)果顯示,5種星蟲動物線粒體13個蛋白質(zhì)編碼基因的Ka/Ks比值均低于1 (0.058 2—0.726 6),表明星蟲動物中蛋白質(zhì)編碼基因遺傳進化中存在一定的純化(負)選擇(表2)。其中,Ka/Ks比值最高的是nad2基因(0.726 6),nad2基因的Ka和Ks比較接近,表明nad2基因在遺傳進化中大多數(shù)為無害的中性選擇,承受的自然選擇壓力較弱。其次是atp8基因,Ka/Ks比值為0.577 3,表明atp8基因承受一定的選擇壓力。cox1、cox3和cob等3個基因的Ka/Ks比值最低(均小于0.1),表明這3個蛋白質(zhì)編碼基因承受強烈的自然選擇壓力,受基因編碼的蛋白質(zhì)功能束縛,從而保證cox1、cox3和cob基因編碼的蛋白質(zhì)生物功能正常,意味著這3個基因?qū)π窍x動物生存進化有重要作用。

表1 星蟲動物線粒體基因組基因變異位點分析
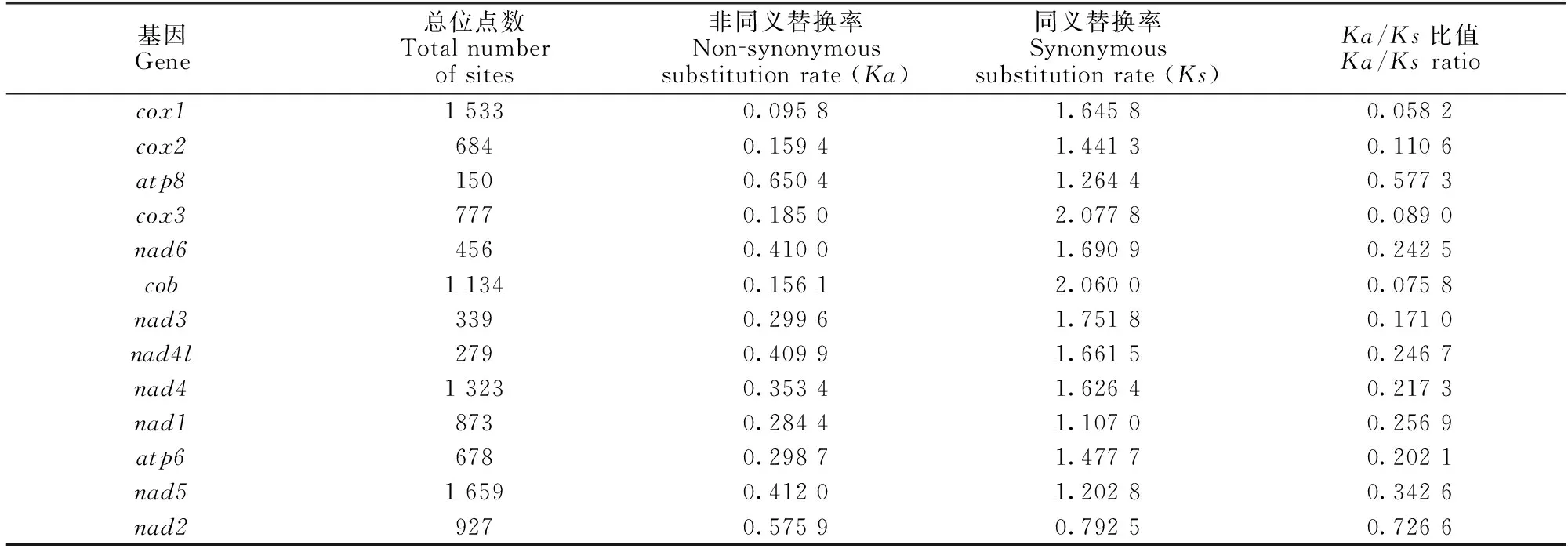
表2 星蟲動物線粒體基因組蛋白質(zhì)編碼基因進化速率分析
2.3 星蟲動物門系統(tǒng)進化分析
采用PhyML 3.0軟件估算,構(gòu)建星蟲動物系統(tǒng)進化樹的最佳進化模型為GTR+G+I。以刺胞動物為外群,星蟲動物及其他8個門共35個線粒體基因組的13個蛋白質(zhì)編碼基因的核苷酸序列為原始數(shù)據(jù),利用該進化模型構(gòu)建系統(tǒng)進化樹(圖6)。研究結(jié)果顯示,不同地理種群的裸體方格星蟲與不同地理種群的可口革囊星蟲首先聚成一支,然后革囊星蟲綱的4種和方格星蟲綱的2種聚合成平行進化的兩個小分支,接著再形成門獨立的進化分支。該結(jié)果符合星蟲動物分類上的親緣關(guān)系,并與傳統(tǒng)的形態(tài)學鑒定分類結(jié)果一致。由圖6可見,星蟲動物與環(huán)節(jié)動物、螠蟲動物聚合成一支大進化分支,軟體動物則是單獨聚合成另一支獨立的平行進化分支,表明星蟲動物與環(huán)節(jié)動物存在近緣關(guān)系,而不是與軟體動物或腕足動物近緣。
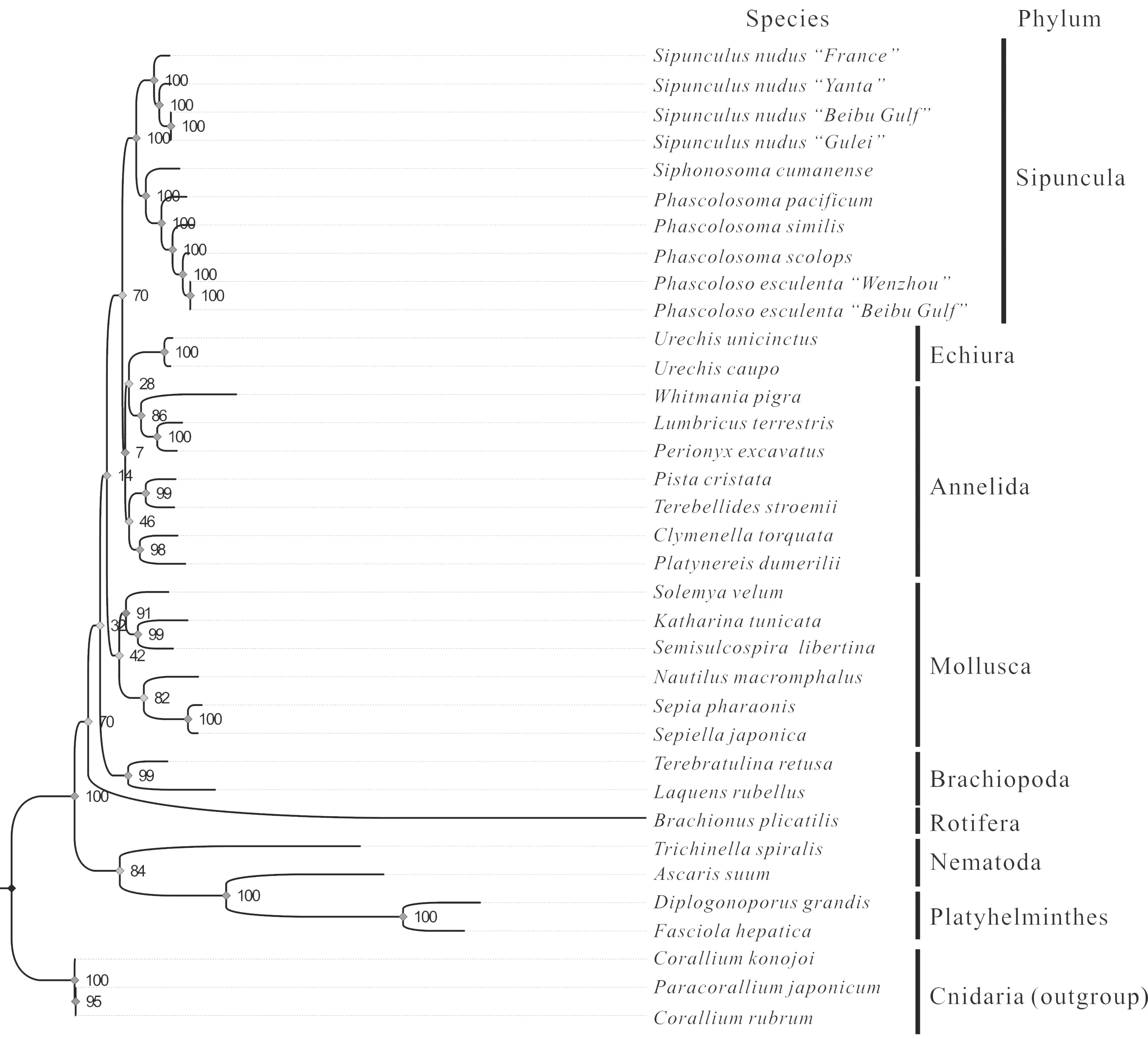
圖6 基于線粒體基因組構(gòu)建的星蟲動物系統(tǒng)進化樹
3 討論
3.1 星蟲動物線粒體基因組的基因序列特征
本研究結(jié)果顯示,廣西北部灣5種常見星蟲動物線粒體基因組長度為15—17 kb,符合無脊椎動物線粒體基因組的長度特征,其長度的差異主要是由非編碼的控制區(qū)長度變異導致的[21]。大多數(shù)后生動物線粒體基因組的堿基組成有偏好性,一般都是富含A、T堿基[22],本研究中的星蟲動物線粒體基因組的堿基組成也呈A、T偏好,其A、T堿基含量為57.5%—65.6%,并且可口革囊星蟲屬的A、T堿基含量普遍高于方格星蟲屬,可能與可口革囊星蟲屬線粒體基因組中含有比例較高的富含A、T堿基的編碼控制區(qū)有關(guān),是否與可口革囊星蟲屬耐受低氧的進化選擇有關(guān)尚需進一步的研究分析。無脊椎動物線粒體典型基因組成包括13個蛋白質(zhì)編碼基因,22個轉(zhuǎn)運RNA基因和2個核糖體RNA基因,以及一些非編碼區(qū)域,并且線粒體基因組的基因排列在長期進化過程中一般是保持穩(wěn)定的,特別是蛋白質(zhì)編碼基因[22],本研究的結(jié)果類似。5種星蟲動物的線粒體基因均編碼在H鏈上,13個蛋白質(zhì)編碼基因排列高度保守,存在3個高度保守的基因排列區(qū)塊,尚未發(fā)現(xiàn)蛋白質(zhì)編碼基因有易位的進化現(xiàn)象,僅在管體星蟲屬和口革囊星蟲屬星蟲動物中發(fā)現(xiàn)tRNAMet基因重排進化。在環(huán)節(jié)動物和螠蟲動物也有類似的現(xiàn)象報道[13,23],環(huán)節(jié)動物和螠蟲動物的線粒體基因組基因組成和排列在進化過程中高度保守,進化過程中可能存在“棘齒”現(xiàn)象,導致環(huán)節(jié)動物和螠蟲動物的線粒體所有的基因均在同一條鏈上編碼,并且環(huán)節(jié)動物線粒體基因組存在6個保守的基因區(qū)塊,螠蟲動物線粒體基因組存在4個保守的基因區(qū)塊,它們的線粒體基因排列僅有部分tRNAs基因發(fā)生易位,可能與其相對原始的進化地位有關(guān)。星蟲動物較為原始的進化地位可能導致其線粒體基因組的基因重排頻率低于其他高等無脊椎動物,尚需進一步的研究證實。
3.2 星蟲動物線粒體基因組的遺傳變異
線粒體基因組的基因由于具備遺傳變異信息豐富、序列進化相對保守和母系遺傳等優(yōu)點成為后生動物的起源、進化、系統(tǒng)發(fā)育關(guān)系和群體遺傳多樣性研究的理想分子標記[24,25]。一般而言,進化速率較慢的基因適合種、屬、科等高級分類階元之間系統(tǒng)進化分析。而同一物種內(nèi),不同種群間的遺傳多樣性分析則需要選擇進化速率較快、遺傳變異水平較高基因作為分子標記。本研究結(jié)果表明,在5種星蟲動物中cox1、cox2和cob等3個基因進化速率最慢、遺傳變異水平最低,適合作為星蟲動物種屬系統(tǒng)進化研究以及不同種間生物條形碼構(gòu)建的分子標記。nad6、nad4、nad5和nad2等4個基因的遺傳變異水平比較高(大于60%),變異位點數(shù)量較多,適合作為星蟲動物種群遺傳多樣性研究的分子標記。此外,線粒體基因組基因的進化速率一般受純化(負)選擇、突變和定向(正)選擇的控制,非同義替換率和同義替換率的比值(Ka/Ks)是確定線粒體基因組蛋白質(zhì)編碼基因進化選擇壓力的一個重要指標[25]。本研究結(jié)果顯示,星蟲動物線粒體13個蛋白質(zhì)編碼基因的Ka/Ks均低于1(0.058 2—0.726 6),表明星蟲動物線粒體的蛋白質(zhì)編碼在基因遺傳進化中存在一定的純化(負)選擇。其中,cox1、cox3和cob等3個基因的Ka/Ks最低(均低于0.1),表明在星蟲動物線粒體的遺傳進化中,這3種蛋白質(zhì)編碼基因承受強烈的自然選擇壓力和功能束縛。在環(huán)節(jié)動物和節(jié)肢動物等無脊椎動物中也有研究報道指出[13],線粒體基因組中細胞色素氧化酶亞基基因(cox1、cox2、cox3和cob)的Ka/Ks最低,承受最強的自然選擇壓力和功能束縛;腺苷三磷酸酶亞基atp8基因的Ka/Ks最高,承受最小的自然選擇壓力和功能束縛[26],甚至在某些軟體動物中線粒體的atp8基因發(fā)生進化丟失現(xiàn)象[27]。而在星蟲動物線粒體中,nad2基因的Ka/Ks為0.726 6,表明星蟲動物線粒體中nad2基因承受最弱的自然選擇壓力和功能束縛,其次才是atp8基因(Ka/Ks為0.577 3)。該研究結(jié)果與環(huán)節(jié)動物和節(jié)肢動物研究結(jié)果稍有不同,是否與星蟲動物耗能較低的穴居生存方式有關(guān),尚需更多的研究分析來證實。
3.3 星蟲動物的系統(tǒng)進化
星蟲動物的分類地位至今還存在不少爭議,因為星蟲動物和軟體動物具有相似的胚胎發(fā)育過程和擔輪幼蟲發(fā)育期,不少研究者認為星蟲動物和軟體動物具有較近的親緣關(guān)系,再加上缺乏環(huán)節(jié)動物的典型特征(身體分節(jié)和具有幾丁質(zhì)剛毛),星蟲動物通常被認為與環(huán)節(jié)動物親緣關(guān)系較遠[8]。然而,本研究中,基于線粒體全基因蛋白質(zhì)編碼基因構(gòu)建的系統(tǒng)進化樹顯示,星蟲動物與環(huán)節(jié)動物的分類地位和親緣關(guān)系較近,與軟體動物的親緣關(guān)系較遠,這與Shen等[13]的研究結(jié)果一致。有研究顯示,星蟲動物和環(huán)節(jié)動物可能起源于分節(jié)的共同祖先。雖然星蟲動幼體和成體都不分節(jié),但是其腦神經(jīng)具有分節(jié)的特征,表明星蟲動物可能由有分節(jié)的祖先進化而來,但在其長期穴居生存的進化過程中分節(jié)的特征逐漸退化,僅在神經(jīng)系統(tǒng)中保持分節(jié)的特征[1]。此外,星蟲動物的線粒體基因組基因排列與環(huán)節(jié)動物的更為相似,星蟲動物的基因排列可能是從環(huán)節(jié)動物基因排列重組衍生而來,兩者具有相似的保守基因排列區(qū)塊,僅部分tRNAs基因發(fā)生易位;而軟體動物線粒體基因組進化重組率較高,基因排列高度異化,與星蟲動物和環(huán)節(jié)動物明顯不同[27]。該結(jié)果亦說明星蟲動物和環(huán)節(jié)動物的親緣關(guān)系較近,而與軟體動物的親緣關(guān)系較遠。當然,這一結(jié)論的證據(jù)尚不夠充分,為了深入研究星蟲動物的系統(tǒng)進化關(guān)系,尚需要挖掘更多其他星蟲動物種類的線粒體基因組數(shù)據(jù)以及核基因數(shù)據(jù)來聯(lián)合研究分析。
4 結(jié)論
本研究分析了廣西北部灣5種常見星蟲動物線粒體基因組序列信息,研究結(jié)果顯示,星蟲動物線粒體基因組具有典型的無脊椎動物線粒體基因組的特征,其編碼基因包括13個蛋白質(zhì)編碼基因,22個轉(zhuǎn)運RNA基因和2個核糖體RNA基因。星蟲動物線粒體基因組的基因均編碼在H鏈上,存在3個高度保守的基因排列區(qū)塊,特別是其13個蛋白質(zhì)編碼基因排列高度保守,尚未發(fā)現(xiàn)蛋白質(zhì)編碼基因有易位的進化現(xiàn)象,僅在管體星蟲屬和口革囊星蟲屬星蟲動物中發(fā)現(xiàn)tRNAMet基因重排進化,與環(huán)節(jié)動物和螠蟲動物線粒體基因組特征較為相似,表明星蟲動物進化地位較為原始。在星蟲動物中,cox1、cox2和cob等3個基因進化速率最慢、遺傳變異水平最低,適合作為其種屬系統(tǒng)進化研究以及不同種間生物條形碼構(gòu)建的分子標記;nad6、nad4、nad5和nad2等4個基因的遺傳變異水平比較高,變異位點數(shù)量較多,適合作為其種群遺傳多樣性研究的分子標記。星蟲動物線粒體遺傳進化過程存在較強的純化(負)選擇,cox1、cox3和cob等3個基因在星蟲動物線粒體遺傳進化過程承受強烈的自然選擇壓力和功能束縛。系統(tǒng)進化樹研究表明,星蟲動物分為方格星蟲綱和革囊星蟲綱兩個進化分支,星蟲動物與環(huán)節(jié)動物的進化地位、親緣關(guān)系較近,與軟體動物的親緣關(guān)系較遠。