丙戊酸鈉干預(yù)對(duì)癲癇鼠模型海馬區(qū)B1及B2受體表達(dá)的影響
2021-01-15 01:08:56劉恒方劉田田郭世燕
中國(guó)實(shí)用神經(jīng)疾病雜志 2020年24期
劉 方 張 敏 劉恒方 劉田田 郭世燕
1)鄭州大學(xué)第五附屬醫(yī)院,河南 鄭州 450052 2)鄭州大學(xué)第一附屬醫(yī)院,河南 鄭州 450052 3)鄭州新鄭國(guó)際機(jī)場(chǎng)急救中心,河南 鄭州 451161
研究發(fā)現(xiàn)[1-2]B1與B2激肽受體(kinin B1 and B2 receptors;KB1R,KB2R)與癲癇的發(fā)作次數(shù)及發(fā)作持續(xù)時(shí)間密切相關(guān),KB1R激動(dòng)劑促進(jìn)肌糖磷酸化一系列病理變化,誘發(fā)細(xì)胞內(nèi)Ca2+水平升高,繼而導(dǎo)致癲癇的發(fā)作[3-5];此外,KB1R及KB2R可上調(diào)磷脂酶A2表達(dá),使前列腺素增加,從而誘導(dǎo)組織水腫,這些機(jī)制與癲癇發(fā)作后的腦組織水腫有關(guān)。已有學(xué)者[6-7]通過(guò)放射配基結(jié)合技術(shù)探討KB1R及KB2R不同程度阻斷后對(duì)動(dòng)物模型癲癇發(fā)作的影響[8-10],揭示KB1R及KB2R在癲癇發(fā)作過(guò)程中的作用機(jī)制。本研究應(yīng)用丙戊酸鈉干預(yù)癲癇鼠模型,觀察不同時(shí)間內(nèi)海馬區(qū)KB1R及KB2R的表達(dá)變化,探討其作用機(jī)制,試圖尋找癲癇發(fā)作新的作用機(jī)制與治療靶點(diǎn)。
1 資料與方法
1.1實(shí)驗(yàn)動(dòng)物及分組選取鄭州大學(xué)動(dòng)物中心提供的35只健康雄性Wistar成年大鼠,體質(zhì)量(250±15.45)g,月齡3~4個(gè)月,按照隨機(jī)化原則分為癲癇實(shí)驗(yàn)?zāi)P徒M30只,采用氯化鋰-匹羅卡品點(diǎn)燃法制作癲癇模型,再分為癲癇模型干預(yù)組15只,其中癲癇持續(xù)狀態(tài)后急性期組5只(12 h,Ⅰ組),靜止期組5只(5 d,Ⅱ組),慢性期組5只(60 d,Ⅲ組);癲癇模型無(wú)干預(yù)組共15只,也分為3個(gè)小組,急性期無(wú)干預(yù)組5只(12 h,Ⅰa組),靜止期無(wú)干預(yù)組5只(5 d,Ⅱa組),慢性期無(wú)干預(yù)組5只(60 d,Ⅲa組;同時(shí)設(shè)置空白對(duì)照組5只(Ⅳ組)。
1.2氯化鋰-匹羅卡品癲癇模型建立及給藥方法Wistar鼠給予氯化鋰125 mg/kg腹腔注射,18~20 h后給予匹羅卡品10 mg/(kg·次)腹腔注射。觀察鼠模型達(dá)5級(jí)后經(jīng)鼻飼法灌入丙戊酸鈉,每4 h灌胃1次,1 mg/(kg·次)。按照相應(yīng)時(shí)間點(diǎn)處死動(dòng)物,取海馬標(biāo)本置入液氮中保存。
1.3RT-PCR法取約50 mg海馬組織,按相關(guān)文獻(xiàn)法提取總RNA。采用一步反應(yīng)法完成RT-PCR反應(yīng),其引物及產(chǎn)物片斷見(jiàn)表1。

表1 B1、B2受體及內(nèi)參照β-acting基因引物
1.4Westernblot法檢測(cè)KB1R及KB2R蛋白表達(dá)采用美國(guó)Eagle EyeII型凝膠圖像系統(tǒng)分析目標(biāo)蛋白和標(biāo)準(zhǔn)對(duì)照的光密度,取兩者比值作為目標(biāo)蛋白半定量相對(duì)值。

2 結(jié)果
2.1氯化鋰-匹羅卡品癲癇模型的觀察大鼠注射匹羅卡品后,可以觀察到Racine制定的癇性持續(xù)發(fā)作形式。
2.2KB1RmRNA及KB2RmRNA表達(dá)變化給予丙戊酸鈉干預(yù)模型后,與Ⅰa組比較,Ⅰ組KB1R mRNA表達(dá)均顯著下調(diào)(P<0.05),Ⅰ組KB2R mRNA表達(dá)均顯著上調(diào)(P<0.05);與Ⅱa組比較,Ⅱ組KB1R mRNA表達(dá)顯著下調(diào)(P<0.05);與Ⅲa組比較,Ⅲ組KB2R mRNA表達(dá)均顯著上調(diào)(P<0.05);與空白對(duì)照組比較,Ⅰa組及Ⅱa組KB1R及KB2R表達(dá)均顯著上調(diào)(P<0.05),Ⅲa組KB2R表達(dá)均顯著上調(diào)(P<0.05)。見(jiàn)表2。
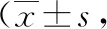
表2 各組B1、B2 激肽受體的mRNA表達(dá)水平比較光密度值OD)
2.3KB1R及KB2R的蛋白質(zhì)表達(dá)變化給予丙戊酸鈉干預(yù)模型后,與Ⅰa組比較,Ⅰ組KB1R蛋白質(zhì)表達(dá)均顯著下調(diào)(P<0.05),Ⅰ組KB2R蛋白質(zhì)表達(dá)均顯著上調(diào)(P<0.05);與Ⅱa組比較,Ⅱ組KB1R蛋白質(zhì)表達(dá)顯著下調(diào)(P<0.05);與Ⅲa組比較,Ⅲ組KB2R蛋白質(zhì)表達(dá)均顯著上調(diào)(P<0.05);與空白對(duì)照組比較,Ⅰa組及Ⅱa組KB1R及KB2R表達(dá)均顯著上調(diào)(P<0.05),Ⅲa組KB2R表達(dá)均顯著上調(diào)(P<0.05)。見(jiàn)表3。
表3 藥物干預(yù)后模型B1、B2 激肽受體蛋白質(zhì)表達(dá)水平比較光密度值OD)
3 討論
分子生物學(xué)技術(shù)研究表明[12-13]KB1R激活后可導(dǎo)致肌糖磷酸化,升高細(xì)胞內(nèi)Ca2+水平;KB1R及KB2R激活后可誘導(dǎo)組織水腫,產(chǎn)生前列腺素[14],細(xì)胞分裂素(絲裂原)活化蛋白激酶可通過(guò)KB1R及KB2R激活,從而導(dǎo)致早期即刻基因的表達(dá)和AP-1易位[15-16]。HOPP等[17]報(bào)道的KB1R的激活加劇了興奮性細(xì)胞毒作用和海馬神經(jīng)元的凋亡。研究發(fā)現(xiàn)癲癇發(fā)作誘導(dǎo)了Wistar鼠海馬的KB1R mRNA重新合成,證實(shí)同期組的KB1R表達(dá)上調(diào)[18-20]。ARGANARAZ等[21-24]用KB1R及KB2R基因敲除小鼠癲癇模型,發(fā)現(xiàn)KB1R基因敲除小鼠自發(fā)癲癇次數(shù)減少,海馬區(qū)細(xì)胞脫失減少,苔蘚纖維增生不明顯;KB1R基因敲除小鼠自發(fā)癲癇次數(shù)增加,海馬區(qū)細(xì)胞脫失明顯增加,苔蘚纖維增生顯著,提示KB1R可能加重顳葉癲癇的病損程度,而KB2R則可能對(duì)顳葉癲癇發(fā)作后的海馬組織有保護(hù)作用[25-27]。
本實(shí)驗(yàn)中氯化鋰-匹羅卡品點(diǎn)燃鼠癲癇模型的KB1R及KB2R mRNA及蛋白質(zhì)表達(dá)水平有較大差異,KB1R在癲癇模型早期表達(dá)上調(diào)較明顯,此后逐漸下降,而且KB1R及KB2R mRNA及蛋白質(zhì)表達(dá)水平基本一致,與國(guó)外研究結(jié)果不完全相符,可能由于本實(shí)驗(yàn)方法較為靈敏的緣故[28-30]。丙戊酸鈉與苯妥英鈉有相同的抑制Na+通道的電生理作用現(xiàn)象,丙戊酸鈉真正的抗癇作用機(jī)制尚不十分清楚[31]。丙戊酸鈉干預(yù)癲癇模型鼠后大鼠海馬組織早期KB1R表達(dá)明顯減少,晚期KB2R表達(dá)明顯增加,支持丙戊酸鈉與顳葉癲癇的KB1R及KB2R表達(dá)變化相。丙戊酸鈉可抑制癲癇早期B1激肽受體的高表達(dá),增加晚期B2激肽受體的表達(dá),本研究也證實(shí)KB1R及KB2R參與了癲癇的發(fā)生與維持機(jī)制,通過(guò)丙戊酸鈉干預(yù)進(jìn)一步表明海馬區(qū)KB1R表達(dá)下調(diào),支持KB1R可能具有細(xì)胞毒作用,KB2R可能有腦細(xì)胞保護(hù)作用。因此,通過(guò)調(diào)節(jié)海馬區(qū)KB1R及KB2R表達(dá)水平亦可能成為抗癲癇新的潛在靶點(diǎn)。
猜你喜歡
童話(huà)王國(guó)·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
作文周刊·小學(xué)二年級(jí)版(2022年20期)2022-05-05 01:33:06
中國(guó)民間療法(2021年5期)2021-06-09 09:21:04
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
創(chuàng)新作文(小學(xué)版)(2019年10期)2019-09-25 08:12:28
飲食科學(xué)(2017年5期)2017-05-20 17:11:53
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年5期)2017-05-04 04:14:38
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
作文與考試·小學(xué)高年級(jí)版(2015年17期)2015-05-30 10:48:04
