木薯蠶絲的13C CP/MAS NMR分析
2021-01-21 14:13:21陳靜黃繼偉彭澤冶寧晚娥
絲綢 2021年1期
關(guān)鍵詞:結(jié)構(gòu)
陳靜 黃繼偉 彭澤冶 寧晚娥
摘要: 木薯蠶絲作為一種極具開發(fā)利用價(jià)值的野蠶絲,其結(jié)晶結(jié)構(gòu)、熱學(xué)性質(zhì)和力學(xué)性能顯著不同于桑蠶絲,對木薯蠶絲二級結(jié)構(gòu)的研究有助于進(jìn)一步認(rèn)識其構(gòu)效關(guān)系,拓展其應(yīng)用領(lǐng)域。13C CP/MAS NMR技術(shù)是研究蠶絲絲素蛋白結(jié)構(gòu)的有效方法之一。文章通過13C CP/MAS NMR測試,分析了木薯蠶絲絲素蛋白的二級結(jié)構(gòu)特征,并將其與桑蠶絲和柞蠶絲進(jìn)行了對比。結(jié)果表明:桑蠶絲中Gly的含量較柞蠶絲和木薯蠶絲高,木薯蠶絲與柞蠶絲相近;Ala的含量及其β-sheet結(jié)構(gòu)較柞蠶絲和木薯蠶絲低,木薯蠶絲最高,三者的α-螺旋結(jié)構(gòu)相差不大;桑蠶絲的不定形區(qū)含量最高,木薯蠶絲最低,與柞蠶絲接近。
關(guān)鍵詞: 木薯蠶絲;桑蠶絲;柞蠶絲;絲素蛋白;二級結(jié)構(gòu);13C CP/MAS NMR
中圖分類號: TS102.333
文獻(xiàn)標(biāo)志碼: A
文章編號: 10017003(2021)01000107
引用頁碼: 011101
DOI: 10.3969/j.issn.1001-7003.2021.01.001(篇序)
13C CP/MAS NMR analysis of cassava silk
CHEN Jing1, HUANG Jiwei1,2, PENG Zeye1, NING Wane1
(1.College of Biological and Chemical Engineering, Guangxi University of Science and Technology, Liuzhou 545006, China;2.College of Textile and Clothing Engineering, Soochow University, Suzhou 215123, China)
Abstract:
Cassava silk as a kind of wild silk with great development and utilization value, is significantly different from mulberry silk in terms of the crystal structure, thermal properties and mechanical properties. The study on the secondary structure of cassava silk can help to further understand its structure-activity relationship and expand its application field. 13C CP/MAS NMR is an effective method to study the structure of silk fibroin. By 13C CP/MAS NMR test, the secondary structure characteristics of cassava silk were analyzed, and then compared with those of mulberry silk and tussah silk. The results showed that the content of Gly in mulberry silk was higher than that in tussah silk and cassava silk, and the content of Gly in tussah silk and cassava silk was close. The content of Ala and β-sheet structure were lower than those of tussah silk and cassava silk, which were the highest in cassava silk. There was no significant difference in α-helix structure among the three. The content of amorphous region in mulberry silk was the highest, and that of cassava silk was the lowest, which was close to tussah silk.
Key words:
cassava silk; mulberry silk; tussah silk; silk fibroin; secondary structure; 13C CP/MAS NMR
木薯蠶絲是由喂食木薯葉的蓖麻蠶吐絲形成,是一種重要的絲制品原料,可被用于生產(chǎn)高檔蠶絲被、絹紡紗及織物等產(chǎn)品[1-5]。木薯蠶絲還是重要的天然蛋白質(zhì)原料來源,具有良好的生物相容性、可控的生物降解性、無毒和低免疫源性等特點(diǎn),在生物醫(yī)用材料、食品和精細(xì)化學(xué)品等領(lǐng)域被廣泛關(guān)注[6-8]。近年來,一系列有關(guān)木薯蠶絲的加工與處理方法被報(bào)道,包括脫膠、整理與改性等[9-12],這些報(bào)道為木薯蠶絲的進(jìn)一步開發(fā)與利用提供了參考。同時(shí),木薯蠶絲結(jié)構(gòu)與性能的研究也逐漸深入。楊瑩瑩等[13]通過掃描電鏡、X射線衍射、紅外光譜和力學(xué)性能測試等手段詳細(xì)對比了木薯蠶絲與桑蠶絲和柞蠶絲之間的差異。結(jié)果表明,木薯蠶絲呈現(xiàn)扁平的帶狀,具有高度的β-折疊結(jié)構(gòu),富含丙氨酸(Ala-Ala-Ala)結(jié)晶單元,木薯蠶絲的斷裂強(qiáng)度和斷裂伸長率均優(yōu)于桑蠶絲,與柞蠶絲相似。黃繼偉等[14]通過微商熱重法、差示掃描量熱法和動態(tài)熱機(jī)械分析法等一系列熱分析手段對木薯蠶絲的熱學(xué)性能進(jìn)行了表征,并與桑蠶絲和柞蠶絲進(jìn)行了對比。結(jié)果表明,木薯蠶絲比桑蠶絲具有更好的熱穩(wěn)定性,略好于柞蠶絲。這些研究進(jìn)一步拓展了人們對木薯蠶絲結(jié)構(gòu)與性能的認(rèn)識,為深度開發(fā)和利用木薯蠶絲提供了數(shù)據(jù)基礎(chǔ)。
核磁是研究蛋白質(zhì)結(jié)構(gòu)的有效方法,隨著固態(tài)交叉極化/魔角旋轉(zhuǎn)(CP/MAS)NMR技術(shù)的發(fā)展,在利用核磁技術(shù)研究蠶絲及其絲素蛋白的結(jié)構(gòu)上取得了顯著的成果[15]。ASAKURA等[16-17]通過對蠶絲絲素紡前和紡后的13C化學(xué)位移進(jìn)行分析,研究發(fā)現(xiàn)蠶絲絲素在受溶劑、機(jī)械處理和拉伸、壓縮等力學(xué)處理時(shí)會導(dǎo)致silk Ⅰ向silk Ⅱ發(fā)生結(jié)構(gòu)轉(zhuǎn)變,且silk Ⅰ到silk Ⅱ的結(jié)構(gòu)轉(zhuǎn)變只發(fā)生在蠶絲絲素蛋白的重復(fù)(AGSGAG)n區(qū),主要表現(xiàn)為α-helix構(gòu)象向β-sheet構(gòu)象的轉(zhuǎn)變;并探究得到絲氨酸(Ser)的OγH基團(tuán)可能參與分子間氫鍵的形成,從而有助于β-sheet的形成,促進(jìn)了silk Ⅱ結(jié)構(gòu)的形成,而酪氨酸(Tyr)則可以有效地破壞β-sheet構(gòu)象的層狀結(jié)構(gòu),導(dǎo)致局部結(jié)構(gòu)無序,不利于silk Ⅱ結(jié)構(gòu)的形成[18-19];進(jìn)一步通過對silk Ⅰ和silk Ⅱ這兩種絲素蛋白的Ala、Ser和Gly殘基的13C CP/MAS NMR化學(xué)位移進(jìn)行分析,制得了化學(xué)位移等值線圖,確定了silk Ⅰ和silk Ⅱ的構(gòu)象[20-21]。這些研究不僅為人們認(rèn)識和了解絲蛋白二級結(jié)構(gòu)提供了參考,更為利用13C CP/MAS NMR技術(shù)研究絲蛋白二級結(jié)構(gòu)提供了方法啟示。同時(shí),這些研究也表明13C CP/MAS NMR方法不僅可以粗略地定量分析蠶絲的主要氨基酸組成,更主要的是可提供蠶絲絲素的構(gòu)象信息。因此,本文利用13C CP/MAS NMR技術(shù)對木薯蠶絲的二級結(jié)構(gòu)進(jìn)行了測試與分析,并將其與桑蠶絲和柞蠶絲進(jìn)行對比,分析三種蠶絲的氨基酸的組成及結(jié)構(gòu)上的差異,以期為進(jìn)一步深入認(rèn)識木薯蠶絲的結(jié)構(gòu)特征提供參考。
1 實(shí) 驗(yàn)
1.1 材 料
2017年春季在柳城縣收購的桑蠶繭(廣西壯族自治區(qū)柳州市柳城鵬鑫源繭絲綢有限公司),2017年春季飼養(yǎng)的柞蠶繭和木薯蠶繭(廣西壯族自治區(qū)蠶業(yè)技術(shù)推廣總站),滲透劑JFC(山東優(yōu)索化工科技有限公司),碳酸鈉、碳酸氫鈉(永華化學(xué)科技(江蘇)有限公司),雙氧水(荊州雙雄化工科技有限公司),上述所用試劑均為分析純。
1.2 儀 器
YB 71旦尼爾電子天平(常州市幸運(yùn)電子設(shè)備有限公司),HH-6數(shù)顯恒溫水浴鍋(國華電器有限公司),Y(B) 802 N八籃恒溫烘箱(溫州大榮紡織儀器有限公司),Agilent-NMR-VNMRS 600型核磁共振儀(美國安捷倫公司)。
1.3 方 法
1.3.1 蠶絲準(zhǔn)備與脫膠處理
1)蠶絲準(zhǔn)備:分別取桑蠶繭、柞蠶繭和木薯蠶繭若干粒,剝?nèi)ダO衣,取出蠶蛹,將蠶繭上的蠶絲撕扯至蓬松狀態(tài),備用。
2)蠶絲脫膠處理:因桑蠶繭絲與柞蠶繭絲、木薯蠶繭絲的表面性狀、雜質(zhì)及含膠量不同,故實(shí)驗(yàn)采用了兩種脫膠工藝,分別對桑蠶繭絲和柞蠶繭絲、木薯蠶繭絲進(jìn)行脫膠。
桑蠶絲脫膠方法:取一定量的桑蠶繭絲放入質(zhì)量分?jǐn)?shù)05%的Na2CO3溶液中(浴比1︰100),在100 ℃恒溫水浴鍋中水浴75 min,取出后在40 ℃的溫水中清洗2~3次,再進(jìn)行第二次脫膠,溶液配比和過程與第一次一致,水浴時(shí)間為60 min,取出脫膠后的蠶絲在去離子水中清洗2~3次。
柞蠶絲及木薯蠶絲脫膠方法:配置1 000 mL脫膠溶液(質(zhì)量分?jǐn)?shù)5%的Na2CO3、質(zhì)量分?jǐn)?shù)0.1%的滲透劑JFC、質(zhì)量分?jǐn)?shù)1%的30%濃度的H2O2)加入燒杯,取一定量的柞蠶絲和木薯蠶絲分別放入溶液中(浴比1︰100),在100 ℃恒溫水浴鍋中水浴45 min后取出,用去離子水清洗2~3次后進(jìn)行二次脫膠,二次脫膠溶液為0.5% NaHCO3溶液,其他配比不變,水浴時(shí)間為60 min。
1.3.2 核磁共振測試
采用600 MHz(14.1T)的Agilent-NMR-VNMRS 600 NMR譜儀,13C的共振頻率150 MHz,在氮化硼“雙軸承轉(zhuǎn)子”中分別測定三種約200 mg以丙酮溶液為溶劑的蠶絲絲素樣品,旋轉(zhuǎn)頻率約5 kHz,探針溫度80 ℃,由TCI三共振反式超低溫探頭維持,90°脈沖寬度4.3 μs,單脈沖序列接觸時(shí)間1 ms,脈沖重復(fù)時(shí)間間隔2 s。總共收集了1 024次掃描,弛豫延遲5 s,所得到的13C CP/MAS NMR圖譜中的化學(xué)位移參考外部金剛烷,從TMS轉(zhuǎn)化為ppm,其光譜寬度為200 ppm。
2 結(jié)果與分析
2.1 蠶絲的13C CP/MAS NMR圖譜
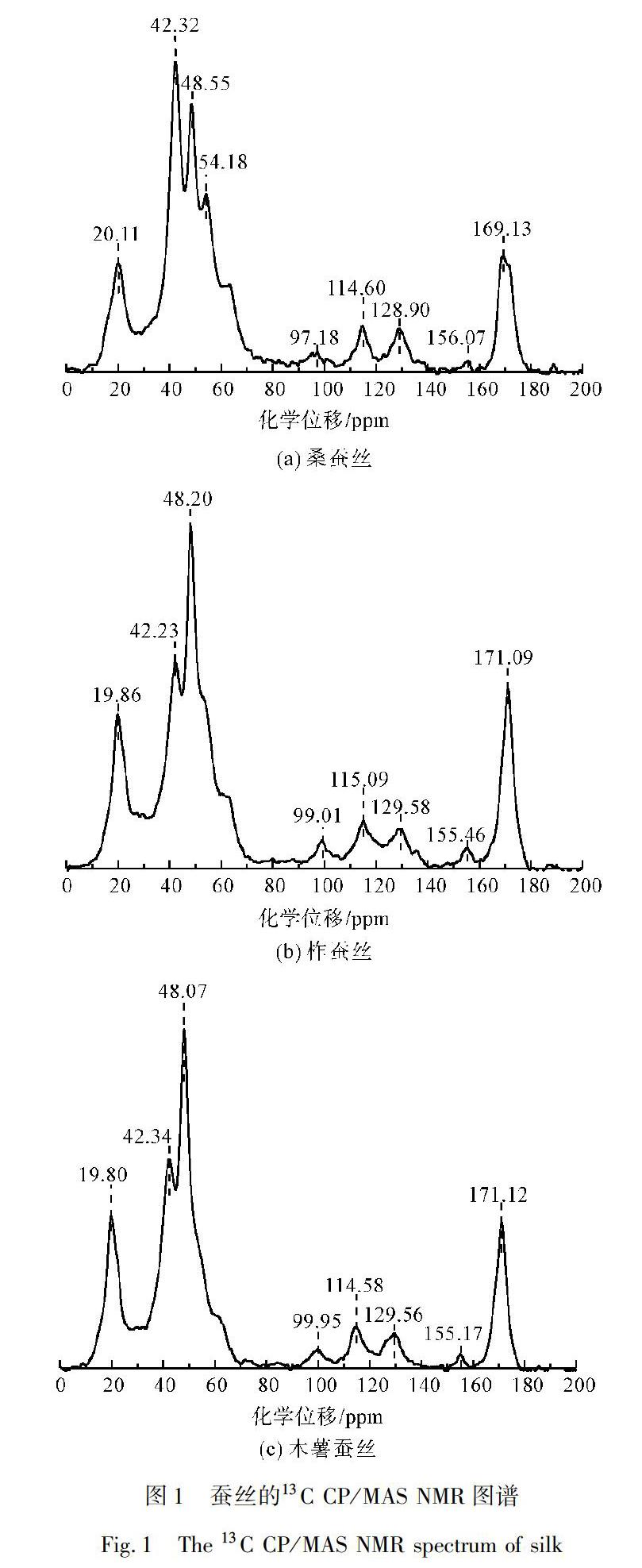
蠶絲絲素蛋白由18種氨基酸組成,其中甘氨酸、丙氨酸、絲氨酸、酪氨酸和天門冬氨酸5種主要氨基酸的總和約占90%[22]。對于不同種類的蠶絲,這5種主要氨基酸的含量又略有不同[23-24]。其中,桑蠶絲的主要氨基酸摩爾比例為甘氨酸約占42.9%、丙氨酸約占30%、絲氨酸約占12.2%、酪氨酸約占4.8%、天門冬氨酸約占1.9%;柞蠶絲的主要氨基酸摩爾比例為甘氨酸約占26.7%、丙氨酸約占48.1%、絲氨酸約占9.1%、酪氨酸約占4.1%、天門冬氨酸約占4.2%;木薯(蓖麻)蠶絲的主要氨基酸摩爾比例為甘氨酸約占33.2%、丙氨酸約占48.4%、絲氨酸約占5.5%、酪氨酸約占4.5%、天門冬氨酸約占2.7%。13C CP/MAS NMR技術(shù)可清晰地分辨出上述5種主要氨基酸中的4種,即甘氨酸、丙氨酸、絲氨酸和酪氨酸,其他氨基酸的峰位通常會被掩蓋,并不容易辨識[25]。
圖1為三種蠶絲的13C CP/MAS NMR圖譜,圖譜中可清晰地辨識出甘氨酸(Gly)、丙氨酸(Ala)、絲氨酸(Ser)和酪氨酸(Tyr)的化學(xué)位移。圖1(a)中,各峰的化學(xué)位移及其對應(yīng)的氨基酸為20.11 ppm(Ala Cβ)、42.32 ppm(Gly Cα)、48.55 ppm(Ala Cα)、54.18 ppm(Ser Cα)、97.18 ppm(SSB)、114.60 ppm(Tyr Cε)、128.90 ppm(Tyr Cγ)、169.13 ppm(Gly CO);圖1(b)
中,各峰化學(xué)位移及其對應(yīng)的氨基酸為19.86 ppm(Ala Cβ)、42.23 ppm(Gly Cα)、48.20 ppm(Ala Cα)、99.01 ppm(SSB)、115.09 ppm(Tyr Cε)、129.58 ppm(Tyr Cγ)、155.46 ppm(Tyr Cξ)、171.09 ppm(Gly CO);圖1(c)中,各峰化學(xué)位移及其對應(yīng)的氨基酸為19.80 ppm(Ala Cβ)、42.34 ppm(Gly Cα)、48.07 ppm(Ala Cα)、99.95 ppm(SSB)、114.58 ppm(Tyr Cε)、129.56 ppm(Tyr Cγ)、155.17(Tyr Cξ)、171.12 ppm(Gly CO)[26-28]。對比三種蠶絲的圖譜可知,桑蠶絲中甘氨酸(42.32 ppm處)的相對強(qiáng)度明顯高于其他氨基酸,而在柞蠶絲和木薯蠶絲中丙氨酸(48.20 ppm處)的相對強(qiáng)度最大,這印證了桑蠶絲中甘氨酸含量最多,而柞蠶絲和木薯蠶絲中丙氨酸的含量最多。為了進(jìn)一步定量分析各氨基酸相對含量及二級結(jié)果的差異,還需要對其進(jìn)行分峰處理。
2.2 譜峰擬合與二級結(jié)構(gòu)分析
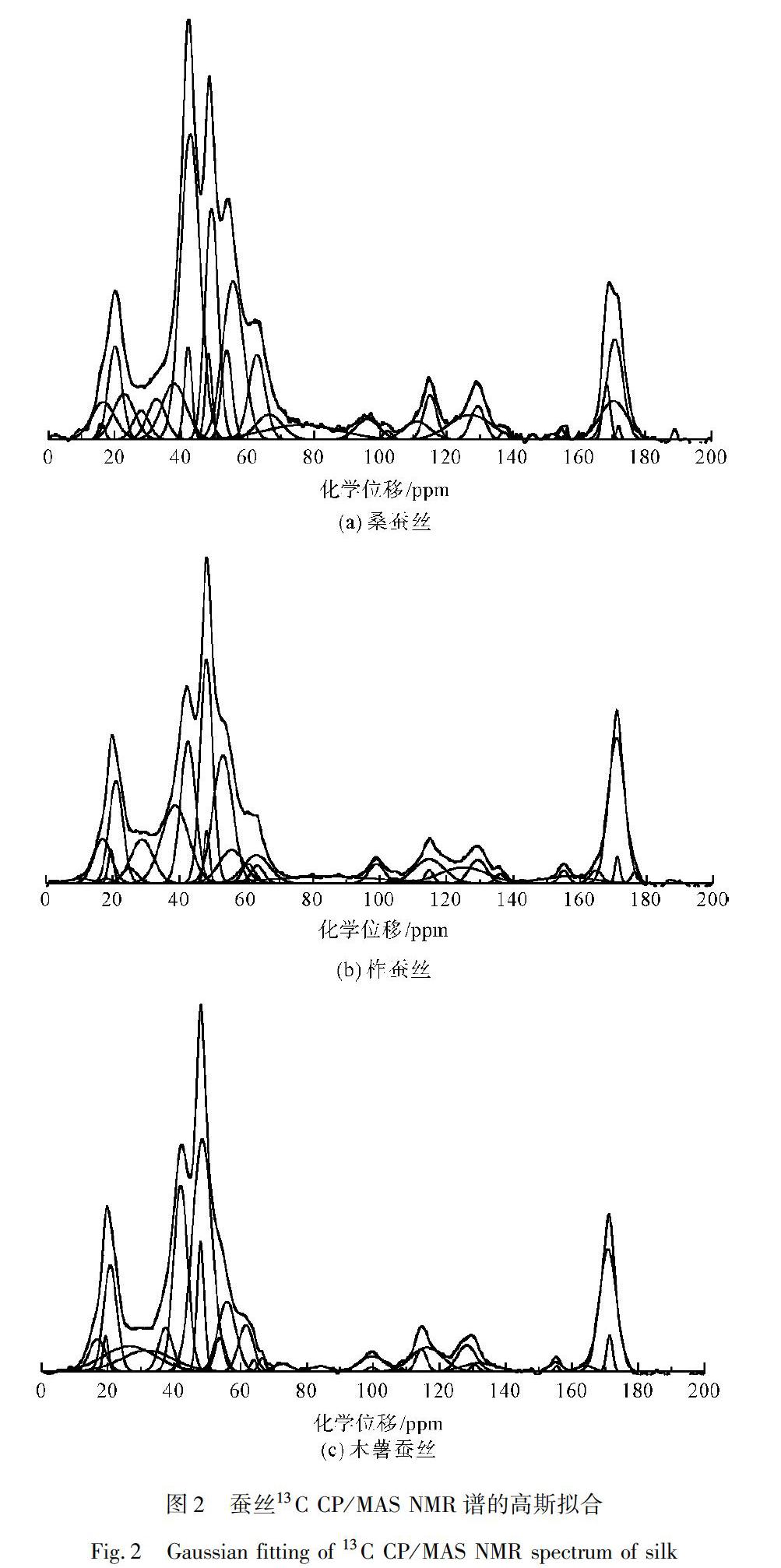
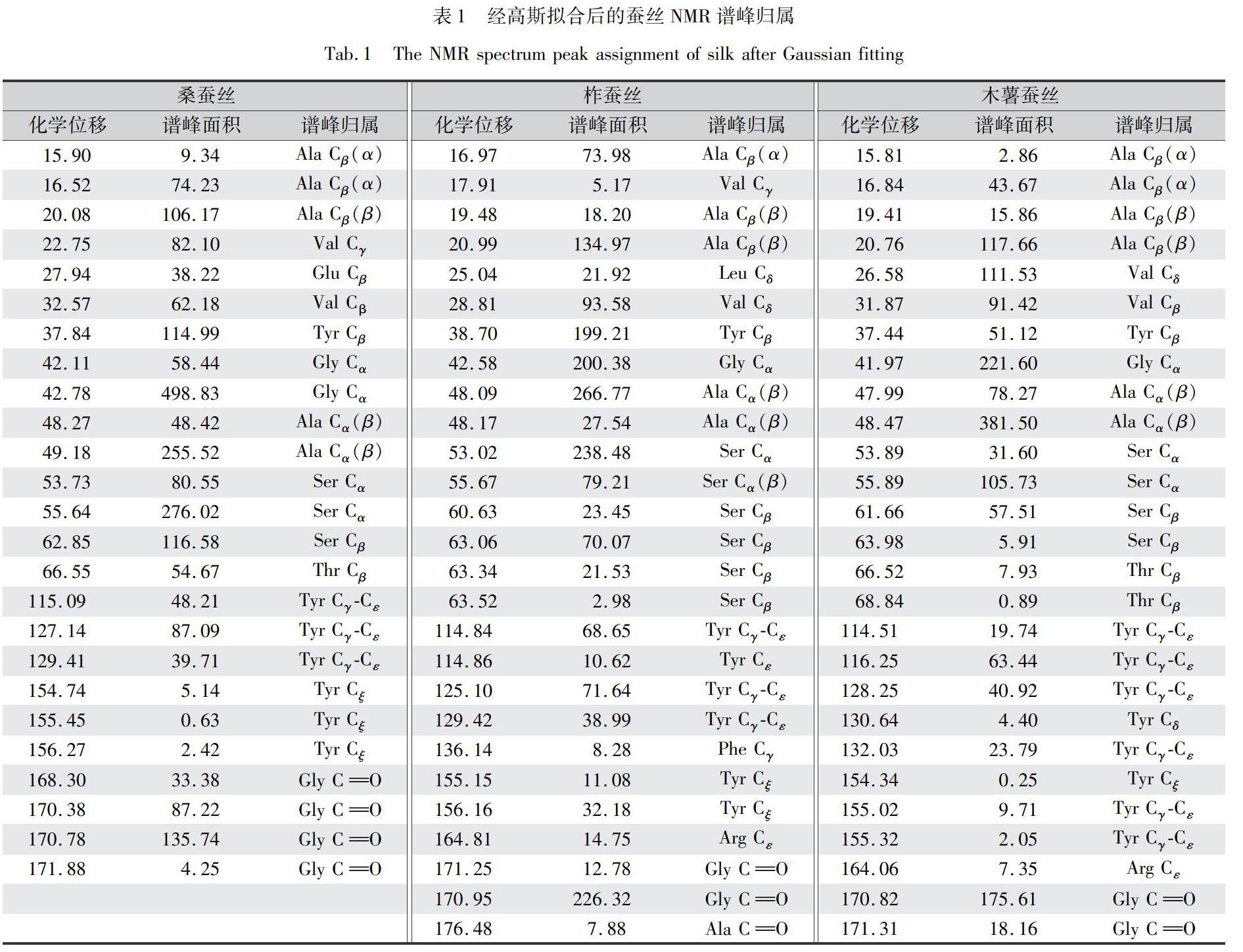
利用Fityk軟件,選用Gaussian線形,對圖1中三種蠶絲的13C CP/MAS NMR譜進(jìn)行分峰擬合處理,結(jié)果如圖2所示。各峰對應(yīng)的化學(xué)位移、譜峰面積和譜峰歸屬,如表1所示。
根據(jù)表1中各氨基酸特征峰對應(yīng)的化學(xué)位移、譜峰面積和譜峰歸屬,對同氨基酸同特征峰類型的面積之和占總譜面積的百分比進(jìn)行分類計(jì)算,結(jié)果如表2所示。
由表2可知,木薯蠶絲中的Gly的相對含量與柞蠶絲接近,但均小于桑蠶絲;就Ala而言,木薯蠶絲的β-sheet結(jié)構(gòu)大于柞蠶絲,而柞蠶絲又大于桑蠶絲,三者的α-helix結(jié)構(gòu)接近。而Ser的相對含量顯示,木薯蠶絲小于柞蠶絲和桑蠶絲。這些結(jié)果說明,木薯蠶絲和柞蠶絲以Ala為主形成結(jié)晶結(jié)構(gòu),而桑蠶絲則以Gly和Ala為主形成結(jié)晶結(jié)構(gòu)。與楊瑩瑩等[13]、王晶等[29]的研究情況相符,即木薯蠶絲和柞蠶絲絲素的結(jié)晶區(qū)主要以(Ala—Ala—Ala)肽鏈結(jié)構(gòu)存在,桑蠶絲絲素的結(jié)晶區(qū)主要以(Ala—Gly—Ala—Gly)肽鏈結(jié)構(gòu)存在。
根據(jù)表1各氨基酸特征峰對應(yīng)的化學(xué)位移、譜峰面積和譜峰歸屬,以譜峰區(qū)域劃分[30-31],計(jì)算各譜峰區(qū)面積占總譜面積的百分比,結(jié)果如表3所示。
由表3可知,在譜峰區(qū)15~30 ppm,柞蠶絲和木薯蠶絲的相對強(qiáng)度比桑蠶絲高,柞蠶絲和木薯蠶絲相近;譜峰區(qū)在30~48 ppm,桑蠶絲的相對強(qiáng)度比柞蠶絲和木薯蠶絲高,柞蠶絲和木薯蠶絲相近。根據(jù)文獻(xiàn)[32]可知,在15~30 ppm譜峰區(qū)主要分布著Ala,在30~48 ppm譜峰區(qū)主要分布著Gly,這進(jìn)一步說明了柞蠶絲和木薯蠶絲的Ala含量較桑蠶絲高,桑蠶絲的Gly含量較柞蠶絲和木薯蠶絲高。
根據(jù)文獻(xiàn)[30],本文對各蠶絲中主要氨基酸特征峰在各譜峰區(qū)所占的比例進(jìn)行計(jì)算,定量比較蠶絲的各主要氨基酸碳峰,用以說明不同氨基酸二級結(jié)構(gòu)在其對應(yīng)的譜峰區(qū)中的比例,如表4所示。
由表4可知,在16~18 ppm譜峰區(qū)內(nèi),主要為Ala的α-helix構(gòu)象,在20~30、48~53 ppm譜峰區(qū)內(nèi)主要為Ala的β-sheet構(gòu)象,三種蠶絲在16~18 ppm譜峰區(qū)內(nèi)的Ala的α-helix構(gòu)象的含量相近,柞蠶絲和木薯蠶絲兩者在20~30、48~53 ppm譜峰區(qū)內(nèi)的Ala的β-sheet構(gòu)象的含量較桑蠶絲高。另外,40~43 ppm譜峰區(qū)內(nèi)三種蠶絲均只有一個Gly Cα峰,說明該區(qū)域幾乎全部來源于Gly Cα的貢獻(xiàn),即可認(rèn)為接近100%。
蠶絲屬于典型的半結(jié)晶聚合物,存在結(jié)晶區(qū)和不定形區(qū),其中結(jié)晶區(qū)主要由反平行β-sheet結(jié)構(gòu)構(gòu)成,不定形區(qū)是由無規(guī)則線圈和松散螺旋結(jié)構(gòu)組成[33-34]。通過三種蠶絲的13C CP/MAS NMR譜圖及分峰擬合結(jié)果可知,Ala呈現(xiàn)出典型的Cα和Cβ構(gòu)象,分別代表了Ala在無定形中的構(gòu)象和結(jié)晶區(qū)中的構(gòu)象。進(jìn)一步地,韓秀文等[30]認(rèn)為,Ala在不定形區(qū)和結(jié)晶區(qū)的含量是不同的(如桑蠶絲Ala結(jié)晶區(qū)占32%,不定形區(qū)占19%;柞蠶絲和木薯蠶絲在結(jié)晶區(qū)占54%,不定形區(qū)占32%),紀(jì)濤等[31]認(rèn)為采用15~30 ppm譜峰區(qū)中Ala Cβ(α)和Ala Cβ(β)兩者的譜峰面積比及無定形區(qū)和結(jié)晶區(qū)Ala的不同含量,可粗略估算出蠶絲中無定形區(qū)所占的比例,計(jì)算過程簡述如下:
1)若將蠶絲看作只有結(jié)晶區(qū)和無定形區(qū)(即不考慮準(zhǔn)結(jié)晶區(qū)等復(fù)雜的情況),記蠶絲絲素中結(jié)晶區(qū)的積分面積和為SA,無定形區(qū)的積分面積和為SB。
2)將Ala在蠶絲結(jié)晶區(qū)中所占的比例記為a,在無定形區(qū)中所占的比例記為b,則應(yīng)有:
a=SAla Cβ(β)SASA=SAla Cβ(β)a(1)
b=SAla Cβ(α)SBSB=SAla Cβ(α)b(2)
則由式(1)和式(2)可知,無定形區(qū)所占的比例記為ηB:
ηB=SBSA+SB=SAla Cβ(α)bSAla Cβ(β)a+SAla Cβ(α)b(4)
3)這里SAla Cβ(α)、SAla Cβ(β)分別為Ala Cβ(α)和Ala Cβ(β)的面積,由分峰擬合得到;a和b的值由韓秀文等[30]、紀(jì)濤等[31]的文獻(xiàn)給出。經(jīng)計(jì)算,無定形區(qū)含量在桑蠶絲中為57%,在柞蠶絲中為45%,在木薯蠶絲中為43%。
分析可知,木薯蠶絲的無定形區(qū)所占比例與柞蠶絲接近,且大于桑蠶絲,這反映木薯蠶絲與柞蠶絲有相似的結(jié)晶度,且均大于桑蠶絲。這一結(jié)論與楊瑩瑩等[9,13]的研究結(jié)果基本一致,但是因計(jì)算式中a和b的值是估計(jì)值,所以結(jié)果是存在誤差的,作為橫向比較是具有參考意義的。
[11]寧晚娥, 趙樹強(qiáng), 林海濤, 等. 抗菌木薯蠶絲的加香整理對其結(jié)構(gòu)和性能的研究[J]. 絲綢, 2019, 56(8): 6-12.
NING Wane, ZHAO Shuqiang, LIN Haitao, et al. Study on the structure and properties of antibacterial cassava silk by the fragrance treatment[J]. Journal of Silk, 2019, 56(8): 6-12.
[12]趙樹強(qiáng), 寧晚娥, 林海濤, 等. 殼聚糖季銨鹽在木薯蠶絲抗菌整理中的應(yīng)用[J]. 絲綢, 2018, 55(7): 13-20.
ZHAO Shuqiang, NING Wane, LIN Haitao, et al. Application of chitosan quaternary ammonium salt in antibacterial finishing of cassava silk[J]. Journal of Silk, 2018, 55(7): 13-20.
[13]楊瑩瑩, 呂智寧, 田偉, 等. 木薯蠶絲結(jié)構(gòu)與性能表征[J]. 紡織學(xué)報(bào), 2017, 38(6): 1-5.
YANG Yingying, L Zhining, TIAN Wei, et al. Structure and properties of cassava silk[J]. Journal of Textile Research, 2017, 38(6): 1-5.
[14]黃繼偉, 趙樹強(qiáng), 寧晚娥, 等. 木薯蠶絲纖維的熱學(xué)性能研究[J]. 絲綢, 2018, 55(8): 7-11.
HUANG Jiwei, ZHAO Shuqiang, NING Wane, et al. Research on thermal properties of cassava silk fiber[J]. Journal of Silk, 2018, 55(8): 7-11.
[15]鄧益斌, 季丹, 周平. 磁共振技術(shù)在絲素蛋白結(jié)構(gòu)與功能研究中的應(yīng)用[J]. 波譜雜志學(xué), 2008, 25(4): 555-572.
DENG Yibin, JI Dan, ZHOU Ping. Application of magnetic resonance methods in the structural and functional characterization of silk[J]. Journal of Spectrum Magazine, 2008, 25(4): 555-572.
[16]ASAKURA T, SUZUKI Y, NAKAZAWA Y, et al. Silk structure studied with nuclear magnetic resonance[J]. Progress in Nuclear Magnetic Resonance Spectroscopy, 2013, 69: 23-68.
[17]ISHIDA M , ASAKURA T, YOKOI. ACTA M, et al. Solvent-and mechanical-treatment-induced conformational transition of silk fibroins studied by high-resolution solid-state 13C NMR spectroscopy[J]. American Chemical Society, 1988, 16: 1.
[18]ASAKURA T, OHGO K, ISHIDA T, et al. Possible implications of serine and tyrosine residues and intermolecular interactions on the appearance of silk I structure of bombyx mori silk fibroin-derived synthetic peptides: high-resolution 13C cross-polarization/magic-angle spinning NMR study[J]. Biomacromolecules, 2005, 6(1): 468-474.
[19]SATO H, KIZUKA M, NAKAZAWA Y, et al. The influence of ser and tyr residues on the structure of Bombyx mori silk fibroin studied using high-resolution solid-state 13C NMR spectroscopy and C-13 selectively labeled model peptides[J]. Polymer Journal, 2008, 40(3): 184-185.
[20]ASAKURA T, OKUSHITA K, P, WILLIAMSON M, et al. Analysis of the structure of bombyx mori silk fibroin by NMR[J]. Macromolecules, 2015(48): 2345-2357.
[21]YAO J, NAKAZAWA Y, ASAKURA T. Structures of bombyx mori and samia cynthia ricini silk fibroins studied with solid-state NMR[J]. Biomacromolecules, 2004, 5: 680-688.
[22]JENKINS J E, CREAGER M S, LEWIS R V, et al. Quantitative correlation between the protein primary sequences and secondary structures in spider dragline silks[J]. Biomacromolecules, 2010(11): 192-200.
[23]CREAGER M S, JENKINS J E, LEIGH A T Y, et al. Solid-state NMR comparison of various spidersdragline silk fiber[J]. Biomacromolecules, 2010, 11(8): 2039-2043.
[24]ZHAO C, ASAKURA T. Structure of silk studied with NMR[J]. Progress in Nuclear Magnetic Resonance Spectroscopy, 2001(39): 301-352.
[25]KRICHELDORF H R, MULLER D, ZIEGLER K, et al. Secondary structure of peptides. IX./sup ?13C NMR CP/MAS investigation of silk proteins[J]. Polymer Bulletin, 1983, 9(6/7): 284-291.
[26]ASAKURA T, WATANABE Y, ITOH T, et al. NMR of silk fibroin. 3. assignment of carbonyl carbon resonances and their dependence on sequence and conformation in bombyx mori silk fibroin using selective isotopic labeling[J]. Macromolecules, 1984, 17(11): 2421-2426.
[27]ASAKURA T, WATANABE Y, UCHIDA A, et al. NMR of silk fibroin. 2. 13C NMR study of the chain dynamics and solution structure of bombyx mori silk fibroin[J]. Macromolecules, 1984, 17(5): 1075-1081.
[28]ASAKURA T, KASHIBA H , YOSHIMIZU H, et al. NMR of silk fibroin. 8. 13C NMR analysis of the conformation and the conformational transition of philosamia cynthia ricini silk fibroin protein on the basis of bixon-scheraga-lifson theory[J]. Macromolecules, 1988, 21(3): 644-648.
[29]王晶, 胡皆漢, 紀(jì)濤. 柞蠶絲蛋白結(jié)構(gòu)的紅外光譜研究及其特異性原因探討[J]. 四川師范大學(xué)學(xué)報(bào)(自然科學(xué)版), 1990, 4(13): 108-110.
WANG Jing, HU Jiehan, JI Tao. Study on the protein structure of tussah silk by IR and the discussion of tussah silk special properties[J]. Journal of Sichuan Normal University(Natural Science), 1990, 4(13): 108-110.
[30]韓秀文, 郭建民, 胡皆漢, 等. 蠶絲蛋白的NMR研究Ⅱ: 桑、柞蠶絲及其絲素的13C CP-MAS NMR研究[J]. 波譜學(xué)雜志, 1991, 8(3): 245-251.
HAN Xiuwen, GUO Jianmin, HU Jiehan, et al. NMR study of silk proteinⅡ: 13C CP-MAS NMR studies of bombyx mori, antheraea pernyi silkorm cocoon and their fibroin in solid state[J]. Chinese Journal of Magnetic Resonance, 1991, 8(3): 245-251.
[31]紀(jì)濤, 韓秀文, 胡皆漢. 蠶絲蛋白的NMR研究Ⅲ: 天蠶繭層固態(tài)13C CP-MAS NMR研究[J]. 波譜學(xué)雜志, 1991, 8(4): 425-431.
JI Tao, HAN Xiuwen, HU Jiehan. NMR study of silk protein Ⅲ: high resolution 13C NMR studies of antheraea yamamai[J]. Chinese Journal of Magnetic Resonance, 1991, 8(4): 425-431.
[32]ASAKURA T, ENDO M, HIRAYAMA M, et al. Glycerin-Induced conformational changes in bombyx mori silk fibroin film monitored by 13C CP/MAS NMR and 1 H DQMAS NMR[J]. Molecular Sciences, 2016, 17: 1-16.
[33]NAKAZAWA Y, NAKAI T , KAMEDA T , et al. A13C NMR study on the structural change of silk fibroin from samia cynthia ricini[J]. Chemical Physics Letters, 1999, 311: 362-366.
[34]SAITO H , TABETA R, ASAKURA T, et al. High-Resolution 13C NMR study of silk fibroin in the solid state by the cross-polarization-magic angle spinning method. conformational characterization of silk i and silk ii type forms of bombyx mori fibroin by the conformation-dependent 13C chemical shifts[J]. Macromolecules 1984, 17: 1405-1412.
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學(xué)評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學(xué)生數(shù)理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
