miR?5088?5p在子癇前期患者胎盤組織中的表達及其對滋養細胞自噬的影響
2021-01-22 01:06:24殷荷王艷華吳琪瑞張輝高麗娜朱亞飛張慧萍吳凱姜怡鄧
實用醫學雜志 2020年24期
殷荷 王艷華 吳琪瑞 張輝 高麗娜 朱亞飛 張慧萍,吳凱 姜怡鄧
寧夏醫科大學1臨床醫學院,2基礎醫學院(銀川750004);3寧夏醫科大學總醫院(銀川750004);4國家衛生健康委代謝性心血管疾病研究重點實驗室(銀川750004);5寧夏血管損傷與修復研究重點實驗室(銀川750004)
子癇前期(preeclampsia,PE)是一種妊娠期女性出現的高血壓、蛋白尿等多種癥狀并存的母嬰共患疾病,嚴重威脅母嬰健康,其發病機制至今仍不明確且尚無有效的防治措施[1]。因此,探索PE 的發病機制及新的防治手段成為產科醫務工作者面臨的新難題。自噬是一種進化上保守的機制,用來維持細胞的動態平衡[2]。目前,自噬在腫瘤、退行性疾病和心血管疾病的發生發展中備受關注[3];近年來,自噬在PE 過程中的作用也逐漸受到重視[4]。研究[5]證實,胎盤滋養細胞自噬水平的異常與多種妊娠并發癥有關,如PE、胎兒生長受限、早產和流產等。然而,其具體作用機制尚不明確,有待進一步研究。microRNA(miRNA)是指能夠負調控基因表達的單鏈非編碼小分子,在多種疾病中起著重要作用[6-7]。許多研究[8]證實,PE 患者存在多種miRNA 的差異表達,到目前為止,只有少數miRNA的作用被證實。其中,miR?5088?5p 在PE 發生、發展過程中的作用未見報道,仍需進一步探討。因此,本研究以胎盤組織和滋養細胞為研究對象,檢測miR?5088?5p 在PE 患者胎盤組織中的表達及在PE 診斷中的臨床價值;同時,體外缺氧誘導滋養細胞發生自噬,模擬PE 發生發展的病理環境,通過過表達miR?5088?5p,檢測自噬相關蛋白的變化,從而揭示miR?5088?5p 在PE 發病過程中的作用機制,為PE 的早期診斷及治療提供新的策略。
1 材料與方法
1.1 材料HTR?8/SVneo 人源胎盤滋養細胞株(上海,瑞鹿);胎牛血清、RPMI?1640 培養基(美國,Gibco);青鏈霉素、胰蛋白酶消化液(北京,solarbio);miR?5088?5p mimics 及miR?5088?5p NC(上海,吉碼);總蛋白提取試劑盒(上海,貝博);LC3B 兔抗人一抗(美國CST 公司,2775);p62 兔抗人一抗(美國abcam 公司,ab109012);β?actin(美國Santa,sc?47778HRP);CK?7鼠抗人一抗(美國abcam公司,ab9021);總RNA提取試劑盒(北京,天根);逆轉錄試劑盒、熒光定量PCR 試劑盒(日本,Takara);miR?5088?5p、U6 引物(廣州,銳博)。
1.2 方法
1.2.1 研究對象及組織標本選取2013年1月至2015年2月在寧夏國龍婦產醫院分娩的PE 孕婦25 例(PE 組),同期健康妊娠分娩的孕婦25 例(PC 組)。納入標準:妊娠20 周后出現收縮壓≥140 mmHg 和/或舒張壓≥90 mmHg,同時伴有蛋白尿(尿蛋白≥0.3 g/24 h 或尿蛋白肌酐比值≥0.3 或隨機尿檢≥+),且無消耗性疾病。組織標本收集前征得所有孕產婦同意并簽署知情同意書,本實驗經寧夏醫科大學倫理委員會審查批準。
1.2.2 qRT?PCR 檢測胎盤組織miR?5088?5p 的表達根據總RNA 提取試劑盒說明書提取PE 組和PC 組胎盤組織的總RNA,取1 μg 逆轉錄為cDNA。通過熒光定量PCR 儀進行擴增,總體系為20 μL,以U6 作為內參,檢測miR?5088?5p 的表達。PCR擴增程序:采用兩步法,95 ℃預變性30 s;95 ℃變性5 s,60 ℃退火34 s,共45 個循環;熔解曲線95 ℃15 s,60 ℃1 min,95 ℃15 s。使用2?ΔΔCt法統計基因的相對表達量,ΔΔCt=[Ct miR?5088?5p(待測樣本)-Ct U6(待測樣本)-[Ct miR?5088?5p(校正樣本)-Ct U6 校正樣本)]。
1.2.3 免疫熒光檢測胎盤組織LC3B 及p62 的表達胎盤組織按照常規方法制備蠟塊,石蠟切片厚度4 μm。石蠟切片脫蠟至水,用枸櫞酸溶液高溫修復抗原,恢復至室溫后,PBS 洗5 min×3 次;3%H2O2室溫孵育30 min,滅活內源性酶,PBS洗5 min×3次;0.5%TritionX?100室溫孵育8 min,PBS洗5 min×3次;山羊血清室溫孵育60 min,棄去血清,分別將稀釋后的CK?7 一抗與LC3B 或p62 一抗混合,均勻滴加于切片,4 ℃孵育過夜;PBS洗5 min×3次;加入熒光二抗,室溫避光孵育2 h,PBS洗5 min×3次;DAPI染細胞核,PBS洗5 min×3次后,滴加抗熒光猝滅劑封片,快速于激光共聚焦顯微鏡下觀察。
1.2.4 細胞培養HTR?8/SVneo 細胞接種于25 cm2的細胞培養瓶中,給予含10%胎牛血清、100 μg/mL青鏈霉素的RPMI?1640 培養基5 mL,置于37 ℃、5 % CO2培養箱內培養。每2 ~3 d 用0.25 %胰蛋白酶消化傳代1 次,取對數生長期的細胞用于后續實驗。
1.2.5 細胞轉染及分組待培養瓶中細胞密度達60%~70%時,嚴格按Lipofectamine2000 說明書進行轉染操作,將miR?5088?5p mimics 及miR?5088?5p NC 分別轉染至HTR8/Svneo 細胞,各分為2 組進行培養:正常組(nomorxia):在37 ℃、5%CO2的條件下培養;缺氧組(hypoxia):在NAPCO Series 8000WJ培養箱中以37 ℃、1%O2和5%CO2的條件下培養。另外設立空白對照組(未進行轉染操作的自然生長HTR8/Svneo 細胞),各組均重復三次。
1.2.6 Western blot測定LC3B和p62蛋白表達收集各組滋養細胞后提取總蛋白;取30 μg 蛋白樣品于12 %分離膠和4 %濃縮膠進行SDS?PAGE 電泳;將蛋白質濕轉至PVDF 膜;用5 %脫脂牛奶封閉2 h;加入一抗4 ℃孵育過夜;PBST 洗滌10 min×3 次;加入HRP 標記的二抗(均1∶5 000),室溫孵育2 h;PBST 洗滌10 min×3 次;ECL 化學發光顯色,分別以LC3BⅡ/Ⅰ灰度值比值作為LC3B 蛋白的表達量,p62 灰度值與內參β?actin 灰度值比值作為p62 蛋白的表達量,實驗重復3 次。
1.2.7 統計學方法用Prism 6.0 統計軟件進行統計學分析,結果以均數±標準差表示,計量資料兩組對比采用t檢驗,多樣本均數間比較采用One?way ANOVA 檢驗,相關性分析以pearson 相關系數表示,P<0.05 表示差異有統計學意義。
2 結果
2.1 研究對象臨床資料的分析與PC 組比較,PE 組年齡差異無統計學意義(P> 0.05),而孕周、收縮壓、舒張壓、血小板計數及新生兒體重差異均有統計學意義(P<0.05),見表1。
表1 研究對象臨床資料分析Tab.1 Analysis of clinical data of study subjects±s
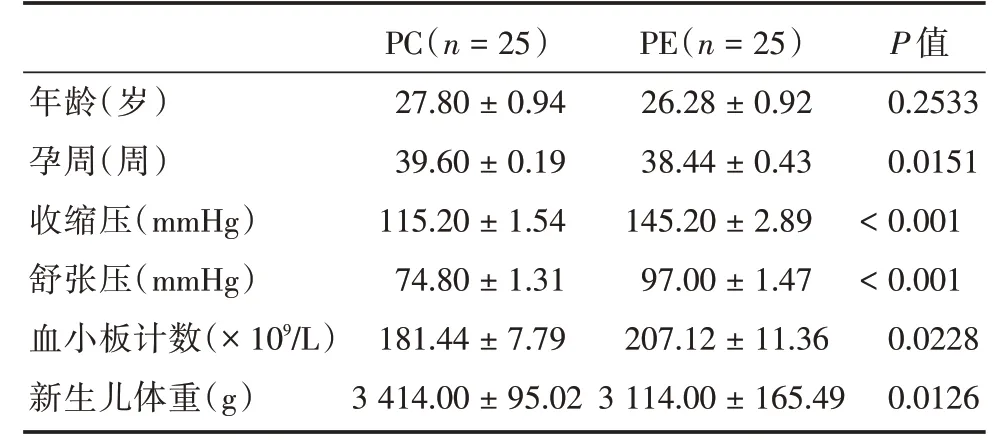
表1 研究對象臨床資料分析Tab.1 Analysis of clinical data of study subjects±s
年齡(歲)孕周(周)收縮壓(mmHg)舒張壓(mmHg)血小板計數(×109/L)新生兒體重(g)PC(n=25)27.80±0.94 39.60±0.19 115.20±1.54 74.80±1.31 181.44±7.79 3 414.00±95.02 PE(n=25)26.28±0.92 38.44±0.43 145.20±2.89 97.00±1.47 207.12±11.36 3 114.00±165.49 P 值0.2533 0.0151<0.001<0.001 0.0228 0.0126
2.2 miR?5088?5p 在胎盤組織中的表達通過qRT?PCR 檢測胎盤組織中miR?5088?5p 表達水平發現,與PC 組比較,PE 組胎盤組織中miR?5088?5p表達明顯下調(P< 0.01),提示miR?5088?5p 可能在PE 發生過程中發揮重要作用,見圖1。
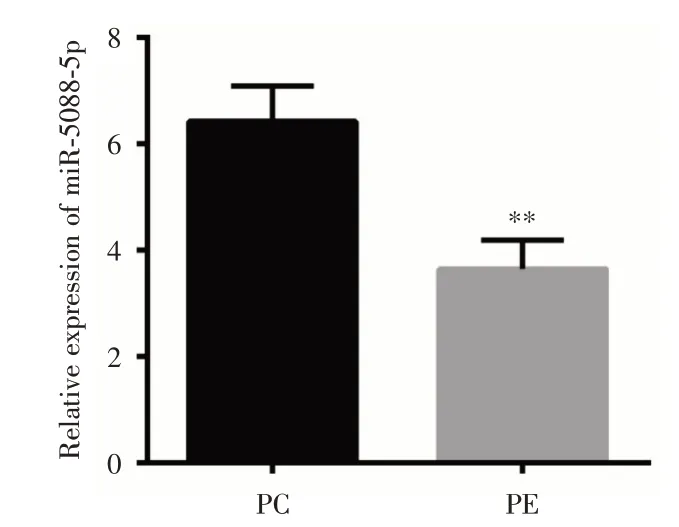
圖1 qRT?PCR 檢測胎盤組織miR?5088?5p 的表達Fig.1 qRT?PCR detection of miR?5088?5p expression in placenta
2.3 miR?5088?5p 表達水平與研究對象臨床特征的相關性分析為了初步探討miR?5088?5p 與PE的關系,進行pearson 相關性分析(見圖2)。miR?5088?5p 的表達水平與收縮壓(r=-0.5 758)、舒張壓(r=-0.6 125)和血小板計數(r=-0.6 082)均呈負相關(P< 0.05);相反,與新生兒體重呈正相關(r= 0.6 458),差異均有統計學意義(P< 0.05),進一步證實miR?5088?5p 在PE 發生發展中的重要作用。
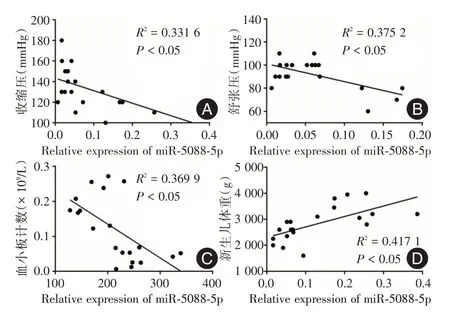
圖2 miR?5088?5p 的表達水平與臨床特征的相關性分析Fig.2 Correlation analysis between the expression level of miR?5088?5p and clinical features
2.4 胎盤組織及胎盤滋養細胞中LC3B 和p62 蛋白的表達胎盤組織免疫熒光結果顯示:與PC 組比較,PE 組LC3B 蛋白表達明顯增加,而p62 蛋白表達減少;同時,體外缺氧干預滋養細胞48 h后,與Nomorxia 組比較,Hypoxia 組胎盤滋養細胞LC3BⅡ/Ⅰ表達增加(P<0.05),而p62 蛋白表達降低(P< 0.05),與組織水平一致,提示在PE 的發病過程中伴隨著自噬的發生,而體外缺氧干預滋養細胞可以模擬PE 發生自噬的病理過程,見圖3。
2.5 過表達miR?5088?5p后胎盤滋養細胞自噬的改變為進一步探討miR?5088?5p的作用,將miR?5088?5p mimics 轉染胎盤滋養細胞,同時分為Nomorxia組和Hypoxia 組,培養48 h 后應用Western blot 檢測各組細胞LC3B 和p62 蛋白的表達。結果顯示:在Nomorxia 組中,轉染miR?5088?5p mimics 后的胎盤滋養細胞LC3BⅡ/Ⅰ表達降低(P<0.001),p62蛋白表達增加(P<0.05),自噬水平降低;同時,Hypoxia組中亦發現相同的自噬水平變化,提示miR?5088?5p可抑制缺氧所致的胎盤滋養細胞發生自噬,從而抑制PE的發生發展,見圖4。
3 討論
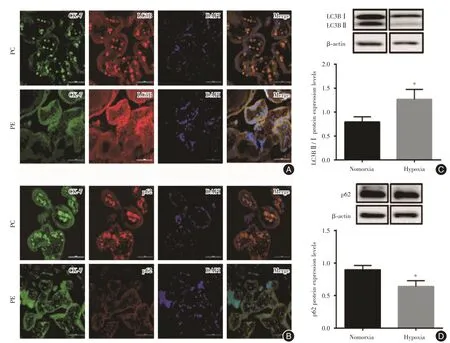
圖3 胎盤組織及胎盤滋養細胞中LC3B 和p62 蛋白的表達Fig.3 Expression of LC3B and p62 proteins in placental tissues and placental trophoblasts
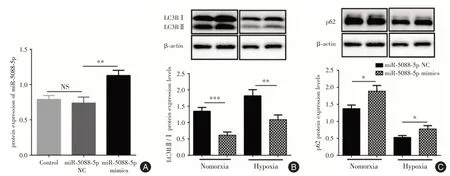
圖4 miR?5088?5p 過表達后檢測其自身和LC3B、p62 蛋白的表達Fig.4 Detection of miR?5088?5p and LC3B,p62 protein expression after overexpression miR?5088?5p
近年來,PE 的發病率和病死率逐年上升[9-10]。然而,PE 的發病機制至今仍不明確。胎盤是母體與胎兒間進行營養物質交換的特有器官,臨床資料顯示,隨著胎兒和胎盤的娩出,PE 的臨床癥狀可迅速恢復正常,可見,胎盤在PE 的發生發展中起著重要作用[11-12]。滋養細胞是胎盤的主要功能細胞,是妊娠維持和穩定的基礎[9]。研究[13]證實,胎盤滋養細胞功能異常與PE 的發展過程密切相關。
自噬廣泛存在于真核細胞中,參與多種細胞的生理病理過程,受多種分子機制調控[14]。研究[15]證實,自噬可在受精、胎盤形成和分娩多個過程中影響妊娠的發生和維持,但目前關于自噬調節PE 發病的具體機制尚不清楚。因此,本研究為了探討自噬在PE 發生發展中的作用,將CK?7 分別與LC3B 和p62 共同定位,胎盤組織免疫熒光檢測發現LC3B 在滋養細胞中的表達增加,而p62 表達減少;同時,通過體外缺氧刺激胎盤滋養細胞,模擬PE 發生微環境,發現與Nomorxia 組比較,Hypox?ia 組胎盤滋養細胞LC3BⅡ/Ⅰ蛋白表達增加,而p62 蛋白表達降低,與PE 胎盤組織中自噬水平變化一致,提示自噬參與了PE 的病理過程,但具體調控機制仍需要進一步研究。
miRNA 是長約22 nt 的小非編碼RNA,在轉錄后水平調節基因的表達變化[16]。同時,miRNA 參與調節各種生理病理過程中的不同信號通路,受到學者的廣泛關注[17]。研究[18-19]表明,孕婦在懷孕期間,受心血管并發癥的影響,各種循環miRNA表達異常。研究[20]發現,miR?133b 在PE 中表達下調,可通過抑制SGK1 促進滋養層細胞的增殖、遷移和侵襲,進而參與PE 的發生發展;提示miRNA可能通過影響滋養細胞的增殖、凋亡、自噬、侵襲和遷移等生物學行為,推動PE 的發生與發展[21]。課題組[22]前期也已發現miR?5088?5p 在缺氧誘導的滋養細胞中表達降低,但是其在PE 中的具體作用仍不清楚。因此,本研究首先通過qRT?PCR 在25 例PC 組和25 例PE 組胎盤組織中檢測miR?5088?5p 的表達,發現miR?5088?5p 在PE 中明顯下調,提示miR?5088?5p 可能在PE 發生過程中發揮重要作用,與以往文獻得出的結論一致;然后,分別將miR?5088?5p 的表達水平與PC 組及PE 組臨床特征進行相關性分析,發現miR?5088?5p 與收縮壓、舒張壓及血小板計數呈負相關,與新生兒體重呈正相關,說明miR?5088?5p 的異常表達與PE 密切相關。同時,為進一步探討miR?5088?5p 在PE 自噬中的作用,體外將miR?5088?5p mimics 轉染滋養細胞,缺氧培養48 h 后,發現過表達miR?5088?5p 后,LC3BⅡ/Ⅰ蛋白表達降低,p62 蛋白表達增高,自噬水平明顯降低。表明miR?5088?5p可以抑制胎盤滋養細胞自噬發生。在PE缺氧模型中轉染miR?5088?5p mimics,發現自噬過程同樣被抑制。這些研究結果提示miR?5088?5p 可以抑制胎盤滋養細胞自噬發生,從而抑制PE 的發生發展。但是,由于標本數量有限,少量的樣本尚不能代表全部的臨床病例,因此,后續將擴大樣本數量進一步驗證,并在血清中探索miR?5088?5p 在PE 疾病中的診斷價值。
綜上所述,研究結果表明miR?5088?5p 在PE中表達降低,并與PE 的臨床特征密切相關,可作為一種保護性因子,抑制PE 胎盤滋養細胞自噬的發生,延緩PE 的發生發展。這為PE 發病機制的研究開拓了一個全新的視角,為研究人員探究PE胎盤滋養細胞自噬過程中miRNA 的調控機制提供了新的思路。并且,由于miR?5088?5p 特殊的生物學功能,它可作為調控滋養細胞自噬的關鍵基因,對其具體機制的進一步研究,將為臨床預防及早期診斷PE 提供重要的理論依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年21期)2018-11-09 01:23:06
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國衛生(2015年9期)2015-11-10 03:11:12
