克氏原螯蝦PcCyc基因克隆及其對光周期的表達響應
2021-01-28 10:35:06蔣琦辰周開亞
水產科學 2021年1期
關鍵詞:分析
謝 偉,蔣琦辰,孫 賓,楊 穎,李 鵬,嚴 潔,周開亞
(1.南京師范大學 生命科學學院,江蘇 南京 210023; 2.江蘇省淡水水產研究所,江蘇 南京 210017 )
周期循環蛋白基因(Cyc基因)是生物鐘基因的核心基因之一。生物鐘是機體內部周期性接近24 h自我維持的時鐘。這個時鐘由分子振蕩器組成,分子振蕩器由轉錄激活因子和時鐘本身及其他基因的抑制劑組成,分別形成正負循環[1]。時鐘控制的生理和行為過程中受環境影響(如光照周期或食物攝入)[2]。Cyc基因是生物鐘環路中的一個重要成員,通常其產物蛋白BMAL1蛋白與CLOCK蛋白形成異源二聚體,結合到period(Per)基因的 E-box盒上激活Per基因轉錄,在時鐘基因調控網絡中起到正調節的作用[3-5]。Cyc基因在脊椎動物中的同源基因被稱為Bmal1基因,Bmal1基因于1997年被克隆獲得,是調節晝夜節律鐘的核心轉錄因子之一。PAS(PER-ARNT-SIM)結構域存在于多種蛋白質中,這些蛋白質在發育和適應環境中起作用[6]。PAS結構域也常見于具有堿性螺旋—環—螺旋(bHLH)結構域的蛋白質中,并且它們成對地充當異二聚體轉錄因子。Cyc基因包含bHLH/PAS轉錄因子[7-8]。bHLH/PAS作為轉錄因子,在中央神經系統的發育中起到至關重要的作用[9-10]。Cyc基因作為重要的生物鐘轉錄基因,有許多重要的生理功能。已有研究發現,Bmal1基因的缺失會導致小鼠完全喪失晝夜節律,并完全破壞體內葡萄糖和甘油三酯水平的周期節律。另外特異性敲除小鼠肝胰腺Bmal1基因,會導致葡萄糖轉運蛋白2的表達喪失晝夜節律性[11],并且小鼠在禁食期間存在低血糖風險[12]。
克氏原螯蝦(Procambarusclarkii),屬節肢動物門、甲殼綱、軟甲亞綱、十足目、爬行亞目、螯蝦科、原螯蝦屬,俗稱小龍蝦,在歐美部分地區也被稱為紅沼澤螯蝦。近些年在中國隨著克氏原螯蝦需求量的增加,克氏原螯蝦的人工繁殖培育不斷發展,養殖面積逐年擴大,克氏原螯蝦已成為中國水產品市場重要的淡水經濟物種[13]。隨著生物技術的發展,時鐘基因的研究已證實,生物體內相關節律鐘基因的紊亂可能導致代謝疾病、癌癥、甚至死亡[14-16]。同時也發現生物的生理行為、新陳代謝、營養獲取等都存在著明顯的節律。認清節律,遵循節律的生活方式可以減少患病的風險[17-18]。在養殖過程中,明確經濟物種的行為、生理、攝食和繁殖等的規律及其分子調控機理可以為健康養殖模式探索提供理論依據。雖然時鐘基因在模式動物中的研究較多,但在水生甲殼動物中的調節機制尚不明確,自甲殼動物發現時鐘基因以來,對其研究多集中于如何保持節律振蕩及對生命活動節律的影響方面[19-21]。筆者克隆克氏原螯蝦Cyc基因并進行生物信息學分析,探究不同光照模式下其mRNA組織差異表達情況,探究Cyc基因的節律振蕩表達模式,以期了解克氏原螯蝦生物鐘基因對其生長發育及代謝的調控作用。
1 材料與方法
1.1 試驗材料與處理
試驗用克氏原螯蝦樣品于徐州(云龍區周邊水域)野外捕捉采集,為除去性別的干擾,在進行試驗時全部選取體長9~11 cm的雄性個體。活體樣本采集完畢后,需要進行光照馴化。在實驗室循環水箱中飼養15 d,溫度恒定25 ℃,24 h充氧,箱底鋪滿沙石,并放置尼龍管和石塊,模擬克氏原螯蝦自然生境躲避模式。用全光譜日光燈模擬室外光照,光暗比12L∶12D(12LD)。馴化15 d之后將樣品分成3組:第1組是馴化15 d之后的樣品直接用于試驗;第2組從中選取健康的雄性個體54只轉入24 h持續黑暗中0L∶24D(24DD);第3組選取健康的雄性個體54尾轉入72 h的持續黑暗中0L∶72D(72DD)。不同光照條件下的克氏原螯蝦均在同一水箱中培養。在不同的光照模式下分別培養結束后,于時間點7:00、11:00、15:00、19:00、23:00、翌日03:00、7:00進行取樣。每個時間點每組取2尾,共取3個平行對照組。使用的試驗操作程序經南京師范大學生物醫學研究倫理委員會[SOXR(江蘇)2015-028]批準。
試驗所用引物名稱及序列見表1,引物合成及測序均由生工生物工程(上海)股份有限公司完成。
1.2 方法
1.2.1 RNA的提取及 cDNA的合成
將克氏原螯蝦的肝胰腺、腦、眼柄組織按照Trizol試劑說明書提取總RNA,用Thermo NanoDrop 2000/2000C(Thermo Fisher Scientific,USA)分光光度計和1.0%的瓊脂糖凝膠電泳檢測RNA的質量和純度,使用反轉錄試劑盒PrimeScriptTMRT Master Mix(TaKaRa,大連)對克氏原螯蝦的RNA進行反轉錄,得到cDNA第一鏈。

表1 試驗所用的引物序列
1.2.2 PcCyc基因表達序列標簽序列的克隆和測序
根據從克氏原螯蝦轉錄組中(Accession code: SRP044128)[22]獲得的PcCyc基因部分序列設計相關引物,引物見表1。將逆轉錄合成的cDNA稀釋10倍作為模板擴增EST序列。反應體系包括:Taq Master Mix 10 μL,ddH2O 7 μL,Cycle-ESF 1 μL,Cycle-ESR 1 μL,cDNA 1 μL。擴增條件為:95 ℃預變性5 min;95 ℃ 30 s,62 ℃ 30 s,72 ℃ 1 min,共29個循環;最后72 ℃ 10 min。PCR產物經1.0%瓊脂糖凝膠電泳檢測,回收后與pMD20-T Vector載體連接,連接產物轉化DH-5α感受態細胞,通過藍白斑篩選挑取陽性克隆菌落,經菌落PCR檢測后于生工生物工程(上海)股份有限公司測序。將測序所得序列在美國國立生物技術信息中心上采用Blast工具進行比對、驗證。
1.2.3 PcCyc基因全長cDNA的克隆
選取已驗證的EST序列,用Primer Premier 5.0軟件設計3′RACE和5′RACE的特異性引物和巢式PCR反應的特異性引物。先對5′RACE PCR進行前處理再進行反轉錄。對提取的總RNA進行CIAP、TAP處理,與5′RACE Adaptor連接后反轉錄合成cDNA。之后進行巢式PCR反應。同樣對3′RACE PCR進行前處理,合成cDNA時要設立M-MLV(-)對照。反應體系:Total RNA(1 μg/μL) 1 μL,3′RACE Adaptor(5 μmol/L) 1 μL,dNTP Mixture(10 mmol/L each) 1 μL,RNase Free deionized H2O 4.5 μL。反應條件:70 ℃,10 min。上述反應結束后立即置于冰上2 min,然后加入下列組分。反應體系:5×M-MLV Buffer 2 μL,RNase Inhibitor (40 U/μL) 0.25 μL,Reverse Transcriptase M-MLV (200 U/μL) 0.25 μL。反應條件:42 ℃ 60 min,70 ℃ 15 min。之后進行巢式PCR反應。將3′RACE和5′RACE的擴增產物經瓊脂糖凝膠電泳,切膠純化并且連接到PMD18-T載體,轉化大腸桿菌(Escherichiacoli)DH-5α感受態細胞,篩選陽性克隆并送生工生物工程(上海)股份有限公司測序。
1.2.4 RACE PCR序列驗證
為驗證RACE PCR擴增的序列是否正確,以EST序列驗證時的cDNA為模板,使用高保真酶,Cycle-QCF/Cycle-QCR 為引物,進行PCR 擴增。反應體系及條件同 EST 序列驗證。反應結束后,取PCR產物于1%的瓊脂糖凝膠上電泳檢測。根據檢測結果確定使用 PCR 產物直接純化還是凝膠回收[使用MiniBEST Agarose Gel DNA Extraction Kit Ver.3.0 (TaKaRa,大連)切膠回收 PCR 產物],用 Cycle-QCF/Cycle-QCR 對 PCR 產物進行測序。
1.2.5 生物信息學分析
利用美國國立生物技術信息中心(http:∥www.ncbi.nlm.nih.gov/BLAST) Blast驗證PcCyc基因的cDNA全長。用ORF Finder查找PcCyc基因的開放閱讀框。在線服務器ExPASy的ProtParam工具分析PcCYC蛋白相關的理化性質。在線軟件Motif Scan可以用來搜索PcCYC蛋白的motif。分別用在線服務器ExPASy的ProtScale工具和在線軟件SignalP分析PcCYC蛋白的疏水性和預測分析信號肽。TMHMM Server v.2.0預測分析PcCYC蛋白跨膜區。在線軟件PSORT Ⅱ Prediction預測分析PcCYC蛋白結構域。PcCYC蛋白二級結構預測使用PSIPRED。三級結構預測分析使用在線服務器Zhang Lab。
1.2.6 不同晝夜節律表達模式分析
克氏原螯蝦不同光照的樣本用Trizol法進行總RNA提取;并各取500 ng用反轉錄試劑盒PrimeScriptTMRT Master Mix(TaKaRa,大連)進行逆轉錄合成cDNA。同時選定克氏原螯蝦β-actin作為內參基因檢測cDNA模板并設計熒光引物,所有的引物見表1。用SYBR?Premix Ex TaqTMⅡ(TaKaRa,大連)試劑在熒光定量儀ABI StepOnePlusTM(Applied Biosystems)上對不同時期的各個組織的基因表達情況進行測定。反應體系:20 μL,包括10 μL 的SYBR?Premix Ex-TaqTMⅡ、0.8 μL 的正反向引物、6.4 μL的PCR反應水、2 μL的cDNA模板和0.4 μL ROX。反應程序:95 ℃ 10 min;95 ℃ 10 s, 60 ℃ 30 s,40 個循環; 95 ℃ 15 s,60 ℃ 1 min, 95 ℃ 15 s。樣本和內參均設置3個重復,反應結束后分析相關數據,采用比較Ct值的方法(基因相對表達量=2-△△Ct)來分析PcCyc基因的相對表達水平。PcCyc基因表達數據的統計分析使用STATISTIC 10.0進行單因子方差分析和Duncan′s多重比較分析,顯著性水平以α=0.05表示。
2 結 果
2.1 cDNA序列及生物信息學分析
5′RACE PCR和3′RACE PCR產物經測序分別得到312 bp的5′端序列,1185 bp和1129 bp的3′端序列。經測序驗證結果表明獲得的RACE PCR產物序列是根據已知EST序列擴增所得(圖1)。PcCyc基因開放閱讀框序列擴增驗證見圖2。

圖1 PcCyc基因RACE PCR序列驗證Fig.1 RACE PCR sequence verification of the PcCyc gene M.DL5000 DNA marker; 1、2.PcCyc 基因. M.DL5000 DNA marker; 1,2.cDNA full-length amplified PCR products.
PcCyc基因的cDNA部分序列分析結果顯示,它具有一個143 bp的5′端非翻譯區(UTR),本試驗中并未獲得3′端非翻譯區,其開放閱讀框包含2010個堿基(GenBank登錄號MN908586),編碼669個氨基酸。PcCyc基因的開放閱讀框中還包含一個典型的bHLH/PAS轉錄因子結構。翻譯過程中起始密碼子為ATG(位于144~146位),終止密碼子為TGA,在本試驗中獲得的序列A+T含量為56.5%,G+C含量為43.5%。

圖2 克氏原螯蝦PcCyc基因cDNA序列及氨基酸序列分析Fig.2 cDNA sequence and amino acid sequence analysis of PcCyc gene in red swamp crayfish P. clarkii N表示糖基化位點; *表示終止密碼子; □表示 CK2; ﹏表示 PKC; 藍色標注表示擴增過程中所用到的引物序列. N denotes Glycosylation site; * denotes termination codon; □ denotes CK2; ﹏ denotes PKC and the primer sequence used in the amplification process is shown in blue.
自美國國立生物技術信息中心下載Cyc基因產物蛋白的同源序列,使用在線軟件Portal CIPES基于最大似然法構建系統發生樹,發現PcCyc基因與甲殼類的Cyc基因聚為一支,且與十足目通訊螯蝦(Pacifastacusleniusculus)的Cyc基因極為相似,氨基酸相似度為93%(圖3),表明本試驗中擴增獲得的PcCyc序列屬于Cyc基因家族。
2.2 PcCYC蛋白性質分析
2.2.1 蛋白質理化特性
PcCYC蛋白的理化性質分析使用在線ProtParam工具進行,對克氏原螯蝦PcCYC蛋白質的理化參數分析顯示,PcCYC蛋白的分子量約為73.963 ku,理論等電點為6.56,原子總數為10 222個,化學分子式為C3192H5035N929O1036S30;在組成PcCYC蛋白的20多種氨基酸中,絲氨酸所占的比例最高,達到13.5%,色氨酸所占的比例最低,為1.2%;脂肪指數為69.07;不穩定指數達到50.54。
2.2.2 PcCYC蛋白功能結構域分析與Motif搜索
PcCYC蛋白結構域的預測分析采用SMART在線軟件進行,分別給出了PcCYC蛋白結構功能域及其相關信息(圖4)。預測結果顯示,PcCYC蛋白含有BHLH/PAS結構域,包括1個HLH結構域和2個PAS結構域。這些結構域可以相互作用形成異二聚體,并結合下游基因的E-Box區域,行使其生物學功能。PcCYC蛋白的Motif搜索利用在線軟件Myhits Motif Scan進行。結果顯示,PcCYC蛋白可能包含19個酪蛋白激酶Ⅱ(CK2)磷酸化位點,14個蛋白激酶C(PKC)磷酸化位點,12個豆蔻酸化位點等(圖2)。
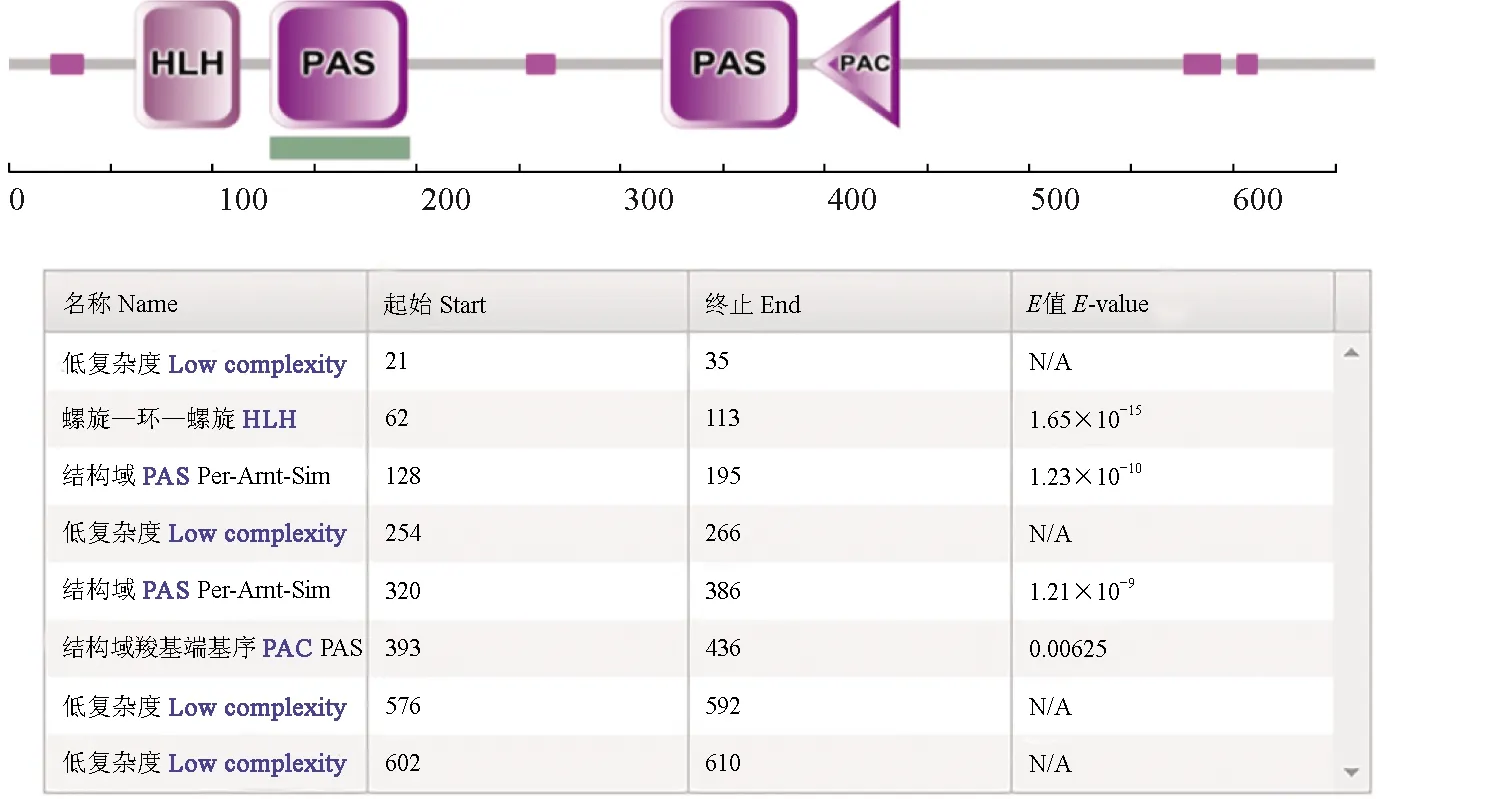
圖4 PcCYC蛋白結構功能域分析預測Fig.4 Analysis and prediction of structure and functional domain of PcCYC protein
2.2.3 PcCYC信號肽預測
SignalP 4.1 Server工具分析PcCYC蛋白并沒有發現信號肽,因此該蛋白為非分泌蛋白,會直接進入細胞質,易于富集,在細胞核中將與PcCLK蛋白形成異二聚體行使其生物學功能。
2.2.4 PcCYC蛋白亞細胞定位分析
采用PSORT Ⅱ Prediction在線服務器對PcCYC蛋白進行亞細胞定位預測。結果顯示,PcCYC蛋白位于細胞核的可能性為95.7%,定位于細胞質的可能性為4.3%,因此PcCYC蛋白為核蛋白,主要存在細胞核中。
2.2.5 PcCYC蛋白空間結構預測
采用PSIPRED在線服務器預測PcCYC蛋白的二級結構,結果顯示,PcCYC蛋白含有11個α-螺旋、12個β-折疊、24個無規則卷曲。采用在線服務器Zhang Lab對PcCYC蛋白進行三維結構預測。圖5中黃色代表β-折疊,藍色代表α-螺旋,紅色代表無規則卷曲;同時PcCYC蛋白含有2個PAS結構域、一個BHLH結構域被分別標出,它們是PcCYC蛋白與其他蛋白互作行使功能的結構域。

圖5 PcCYC蛋白三級結構預測Fig.5 The predicted tertiary structure of PcCYC protein
2.2.6 蛋白質疏水性
采用在線服務器ExPASy的ProtScale工具對PcCYC蛋白的疏水性進行預測,結果顯示,PcCYC蛋白的最高峰值為2.554,最低峰值為-3.578,表明PcCYC蛋白為疏水性蛋白。
2.3 不同光照下PcCyc基因在不同組織中mRNA的表達分析
以β-actin基因作為內參,使用實時熒光定量PCR(qPCR)技術檢測了不同光照條件下PcCyc基因在腦、肝胰腺和眼眪3種組織中的mRNA表達水平,結果表明,3種光照條件下,PcCyc基因在眼柄和肝胰腺中都基本遵循著24 h的節律振蕩,而在大腦中的節律振蕩隨光照周期改變被打破(圖6)。12LD的光照條件下,大腦與眼柄中PcCyc基因基本上有相似的表達規律,但在24DD的光照條件下,相似的規律被打亂了。在24DD光照條件下克氏原螯蝦眼柄中PcCyc基因表達高峰比12LD條件下的提前了4 h,在72DD條件下,節律振蕩恢復,但是與12LD的節律規律略有區別。
3 討 論
生物鐘一直是科學領域研究的熱點問題,它是生命對地球光照以及溫度等環境因子周期變化長期適應而演化的內在自主計時機制[23]。本試驗主要采用RACE PCR技術首次獲得了克氏原螯蝦生物鐘核心基因PcCyc的部分cDNA序列,包括完整的開放閱讀框,共2010個堿基,可編碼669個氨基酸(即PcCYC蛋白)。截至目前,甲殼類Cyc基因,僅有斑點海虱、通訊螯蝦、沙灘跳鉤蝦(Talitrussaltator)和南極磷蝦(Euphausiasuperba)4個物種被報道[8,24-25]。
3.1 克氏原螯蝦PcCyc基因的生物信息學分析
本試驗中使用PcCyc基因翻譯的氨基酸序列對PcCYC蛋白進行相關的理化性質、功能結構域分析。其中功能結構域分析PcCYC蛋白含有典型的bHLH/PAS轉錄因子結構域,在CYC蛋白中該結構域的主要功能是與CLK蛋白結合成異二聚體在時鐘調控網中行使功能[26-27]。對斑點海虱Cyc基因的研究也顯示,其產物蛋白同樣包含BHLH/PAS結構域[8]。這個結構域在果蠅、小鼠、人類等的CYC蛋白中普遍存在[26-28],是CYC蛋白公認的結構域之一,也是CYC蛋白的主要功能結構域。表明了CYC蛋白結構在進化過程中的保守性,也預示了其功能的保守性。已有研究顯示,甲殼動物bHLH-PAS家族的成員通過幼年激素信號傳導調控生長和繁殖[29],通過晝夜節律計時和對氧張力變化的響應來調節運動[30]。這對于養蝦業來說非常重要,水產養殖業經常面臨可能會影響養殖產量的環境條件波動的壓力。

圖6 克氏原螯蝦PcCyc基因mRNA表達水平分析Fig.6 Analysis of mRNA expression levels of PcCyc gene in red swamp crayfish P. clarkii
3.2 克氏原螯蝦PcCyc基因在不同光周期下的組織表達模式分析
Escamilla-Chimal等[31]利用抗體檢測了克氏原螯蝦CLK蛋白在不同光照下不同組織中的表達情況,結果表明,CLK 蛋白在12LD 的光照條件下,眼柄組織中于12:00出現蛋白表達高峰,在24DD的光照條件下,腦組織中于16:00出現蛋白表達高峰,在調控網絡中 CYC 蛋白與CLK 蛋白形成異二聚體共同行使功能,因此推測這兩種蛋白基因可能具有相似的mRNA表達水平。在時鐘基因調控網絡中,時鐘基因蛋白的積累會比mRNA的生成延遲6~8 h[32-34]。本試驗通過實時熒光定量PCR技術檢測PcCyc基因在不同光照下(12LD、24DD和72DD)克氏原螯蝦不同組織(腦、眼柄和肝胰腺)中的mRNA表達水平(圖6)。結果顯示,克氏原螯蝦PcCyc基因在眼柄組織中的mRNA表達水平與之符合,但是腦組織的表達有4 h的出入,原因有待進一步研究。在24DD與72DD條件下,腦組織中該基因的mRNA的表達與 Escamilla-Chimal等[31]的結果相比基本上符合蛋白積累延遲8 h的規律。在克氏原螯蝦肝胰腺中的PcCyc基因,由于肝胰腺屬于外周振蕩器,不同于眼柄中靠近視網膜上的光感受器,肝胰腺中生物鐘基因的節律振蕩主要調節生物的激素水平[35],參與調控機體的生理生化反應振蕩。雖然最終要恢復起初的振蕩水平,但由于生理生化的調節很復雜,可能需要更長的時間去恢復原有的節律振蕩,以使生物適應外界環境的改變。結果分析顯示,在改變光照的條件下,PcCyc基因慢慢地會趨于遵循24 h的晝夜周期振蕩節律,并且在腦組織中有較高的起伏變化,以此證明時鐘基因PcCyc有顯著的內源性自由運轉節律,并受腦組織支配。在斑點海虱腦組織中Cyc基因的mRNA表達雖受到潮汐節律的影響,但還是遵循著近日24 h的節律振蕩,且節律振蕩獨立由腦控制,外部環境條件只會影響節律,而不會控制節律[8]。
4 結 論
在本試驗中,采用RACE PCR技術克隆了克氏原螯蝦的核心生物鐘基因PcCyc的部分cDNA序列(2153個堿基),包括2010個堿基的開放閱讀框(編碼669個氨基酸);同時利用實時熒光定量PCR技術檢測PcCyc基因在不同光照模式下(12LD、24DD和72DD)克氏原螯蝦不同組織(腦、眼柄、肝胰臟)中的差異表達。在改變光照的條件下,PcCyc基因慢慢地會趨于遵循24 h的晝夜周期振蕩節律,在大腦中的節律振蕩隨光照周期改變被打破。試驗結果表明,克氏原螯蝦的時鐘基因PcCyc有顯著的內源性自由運轉節律,并受腦組織支配。本試驗結果為研究克氏原螯蝦生物鐘基因對克氏原螯蝦生長發育及代謝的調控作用奠定了一定的基礎。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
