雷州半島東部近岸海域大型底棲甲殼動物群落結構及其影響因子
2021-02-03 10:18:54陳志劼呂少梁陸麗儀曾嘉維王學鋒
廣東海洋大學學報 2021年1期
關鍵詞:影響
陳志劼,呂少梁,陸麗儀,張 靜,曾嘉維,王學鋒
(1.南方海洋科學與工程廣東省實驗室(湛江),廣東 湛江 524025;2.廣東海洋大學水產學院,廣東 湛江 524088)
預測物種的資源密度及其分布格局,是生態學研究的主要內容之一。作為海洋生態系統的重要組成部分,蝦、蟹、蝦蛄類甲殼動物種類多、數量大,是底棲動物的優勢類群[1],其資源變動對海洋生態系統的穩定會產生重要影響[2-3]。隨著近海漁業資源的衰退和魚類等捕撈對象減少以及人們對海洋產品需求的增加,海洋甲殼動物因繁殖力強、生活史較短等特征在捕撈業和漁業經濟中的地位不斷提高,自20 世紀80 年代以來成為主要的捕撈對象之一[4]。根據1990―2016 年《中國漁業統計年鑒》,甲殼動物海洋捕撈總產量1990 年88.13 萬t,至2016年升至239.63 萬t,整體呈波動上升趨勢[5]。因此,對甲殼動物資源開展定期調查與評估,是了解其資源動態及可持續開發利用的基礎。
雷州半島東部近岸海域潮汐特征復雜,幾乎不受入海河流影響,海水鹽度較穩定,為海洋生物提供了適宜的生境[6-7]。該海域不僅是多種海洋經濟動物產卵、育幼及索餌的場所,也是國家一級保護動物中華白海豚 (Sousa chinensis) 主要棲息場所之一[8-9]。目前,關于雷州半島東部近岸海域生物群落結構研究主要集中在魚類[9-10]和浮游生物[11-12];在甲殼類方面,僅見肖漢洪等[13]硇洲島潮間帶蟹類組成及其分布的研究和張旭豐等[14]蝦拖網副漁獲種類組成的研究。硇洲島海域國家級海洋牧場示范區為本項目研究海域的一部分,研究該海域甲殼動物群落結構及生態因子的調控作用,可為海洋牧場建設效果評估、增殖種類選擇、增殖容量研究等提供基礎數據。
1 材料與方法
1.1 數據來源
2016 年4 月、8 月、11 月和2017 年2 月于雷州半島東部近岸海域(110°30′―110°45′E,20°40′―21°05′N)進行漁業資源底拖網調查,具體站位見圖1。依據《海洋漁業資源調查規范》(SC/T 9403―2012)[15],選取選擇性能小的單船型底拖網作為調查網具,網囊網目長度25 mm。受海況等因素的影響,各站位的實際拖速、拖網時間有偏差,因此在數據分析前先做標準化處理,即統一換算成拖速3 n mile/h 情況下每小時的漁獲量(kg/h)。
采集拖網漁獲物中的甲殼動物樣品,參照文獻[16-18]完成種類鑒定,統計其數量和質量(0.01 g)等。所有站位同步采集環境數據:利用SM-5A 測深儀測量水深(d),YSI 水質參數儀測定溶解氧(DO)、pH、鹽度(S)和水溫(t)。各站位采集500 mL 水樣,利用分光光度法測定葉綠素a(Chl a)濃度。
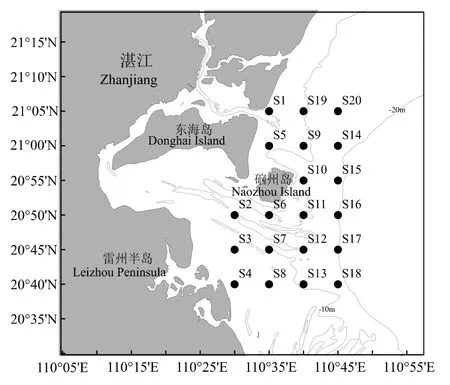
圖1 雷州半島東部近岸海域調查站位Fig.1 Survey stations in the east coastal of Leizhou Peninsula
1.2 數據處理與分析
1.2.1 質量資源密度及環境數據的處理 甲殼動物質量資源密度用掃海面積法計算:
Pi=Ci/(ai q)。
式中,Pi指第i站的甲殼動物資源密度 (質量密度單位:kg/km2);Ci指第i站甲殼動物的每小時漁獲質量(kg/h);ai指第i站拖網的每小時掃海面積(km2/h);q指捕獲率,本研究取0.5,原因有:1)捕撈系數是基于拖網漁具選擇性而得,與漁具規格,海區底質類型、海流等有關;2)為與魚類漁獲物保持一致,增加數據可比性,便于后期對整體漁獲物的比較分析;3)捕獲樣品以小規格蝦蟹類居多。
為平衡群落中優勢種極化作用及偶見種的影響,分析前對數據進行標準化處理。即剔除出現頻率低于10%的種類,四次方根轉換各站位甲殼動物的質量資源密度,環境數據除pH 外做lg (x+1)處理[4,9]。
1.2.2 優勢種分析 利用Pinkas[19]相對重要性指數(IRI)確定甲殼動物優勢種:
IRI=(N+W)×F×10 000,
式中,N為某一甲殼動物的數量比例 (%),W為某一甲殼動物的質量比例 (%),F為某一甲殼動物某站位的出現頻率 (%)。將IRI>1000 的物種定為優勢種。
利用種類更替率 (T) 研究優勢種的季節變化:
T(%)=(a+b-2c)/(a+b-c),
式中,a、b為兩個相鄰季節的優勢種種類數;c為兩個相鄰季節的共有優勢種種類數。
1.2.3 多樣性分析 采用Margalef物種豐富度指數(D)、Shannon-Wiener 多樣性指數 (H′)、Pielou 均勻度指數 (J′) 分析甲殼動物的多樣性,計算公式:
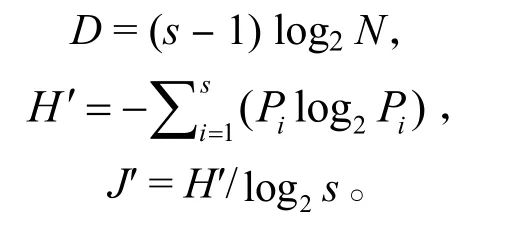
式中,s為甲殼動物種類數,Pi指第i種物種占甲殼動物總物種數的比例,N為甲殼動物總數。
1.2.4 群落結構分析 利用Primer 5.0 軟件進行聚類分析 (Cluster) 和參數多維標序排序 (MDS),分析研究海域甲殼動物的群落聚集性。采用協強系數(Stress) 衡量MDS 二維排序圖結果:當Stress<0.05時,圖形代表性較佳;0.05<Stress<0.1 時,排序較佳;0.1<Stress<0.2 時,圖形有一定解釋意義。
采用單因子相似性分析 (Anosim test) 檢驗甲殼動物群落在不同站位、不同季節間的差異顯著性;采用相似性百分比分析 (Simper test) 計算各季節群落組內相似性和組間相異性的平均貢獻率,并列出貢獻率前五的物種。
1.2.5 群落特征與環境因子的冗余分析 利用R統計軟件[20](Vegan package) 分析甲殼動物群落分布與環境因子的關系。首先,對各站位甲殼動物資源密度矩陣 (站位×各甲殼動物的質量密度) 進行去趨勢對應分析 (DCA),分析結果中,如梯度長度(Lengths of gradient)最大值超過4,則使用單峰模型(CCA)排序;若小于3,則選擇線性模型(RDA)排序;若介于3~ 4,二者均可使用。之后采用蒙特卡洛置換檢驗 (Monte Carlo permutation test) 分析環境因子對甲殼動物分布影響的顯著性。
2 結果與分析
2.1 種類組成
漁獲物樣品共鑒定出甲殼動物98 種 (含未定種1 種),隸屬2 目24 科43 屬(表1)。各季節種類數依次是夏季(74 種)>冬季(62 種)>秋季(59 種)>春季(50 種)。其中,對蝦科 (Penaeidae)和梭子蟹科 (Portunidae) 的種類數較多,分別為23種和27 種,是調查海域甲殼動物的主要類群。四季共有種為變態蟳 (Charybdisvariegata)、斷脊小口蝦 蛄(Oratosquillinainterrupta)、哈氏仿對蝦(Parapenaeopsishardwickii) 等30 種,占甲殼動物種類數的30.6%。
2.2 優勢種
根據相對重要性指數 (IRI) 分析,四季均出現的優勢種為變態蟳、哈氏仿對蝦、須赤蝦(Metapenaeopsis barbata) 等10 種(表2)。優勢種的季節變化顯著(P<0.05),春―夏季、夏―秋季和秋―冬季間優勢種更替率分別為50.6%、29.5%和38.7%。
2.3 生物多樣性
甲殼動物的多樣性指數(H’)、均勻度指數(J’)和豐富度指數(D)均為夏季最高,冬、春季次之,秋季最低(表3)。
2.4 聚類分析
四季甲殼動物群落結構的協強系數 (Stress) 為0.10~0.18,較好地反映了群落間的相似度(圖2)。其中,春季的S2 站位和秋季的S17 站位與同季其他站位相似性較低,暫不納入群落分組。相關原因有待進一步研究。
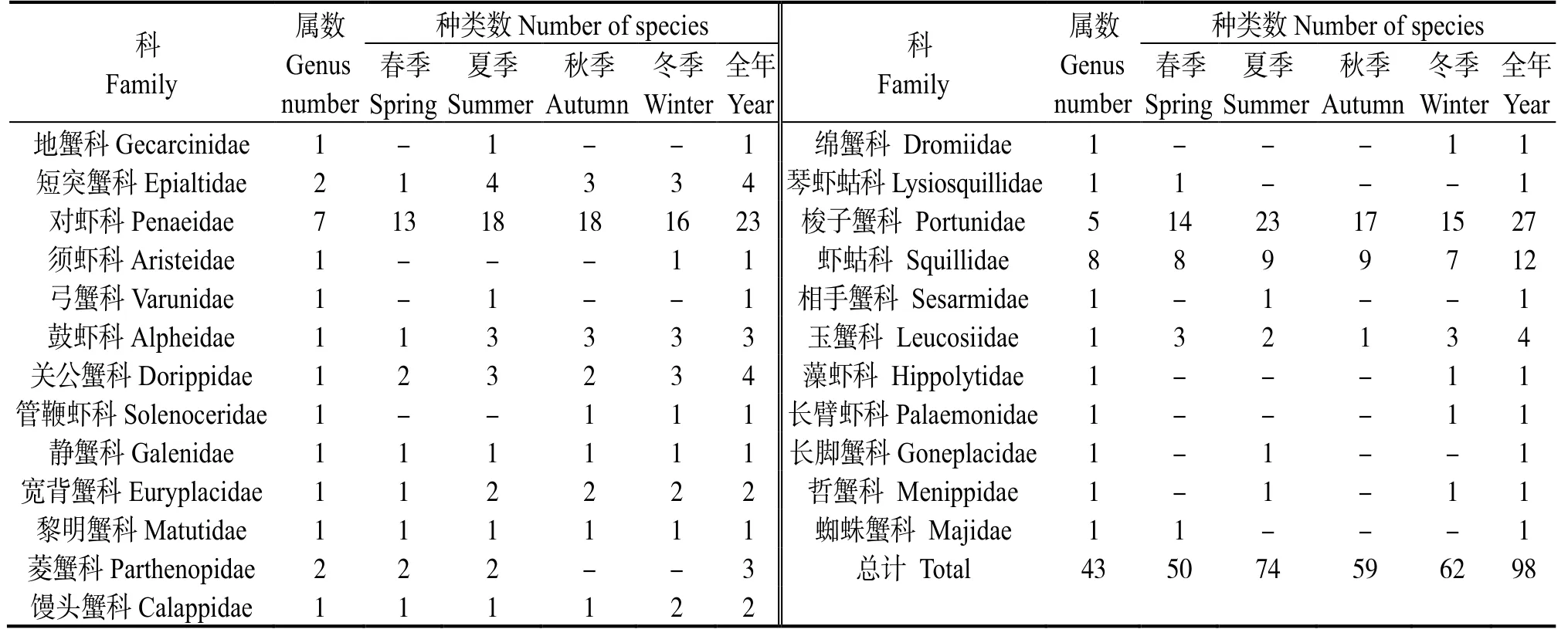
表1 雷州半島東部近岸海域甲殼動物的科、屬和種Table 1 Composition of crustaceans in the east coastal of Leizhou Peninsula
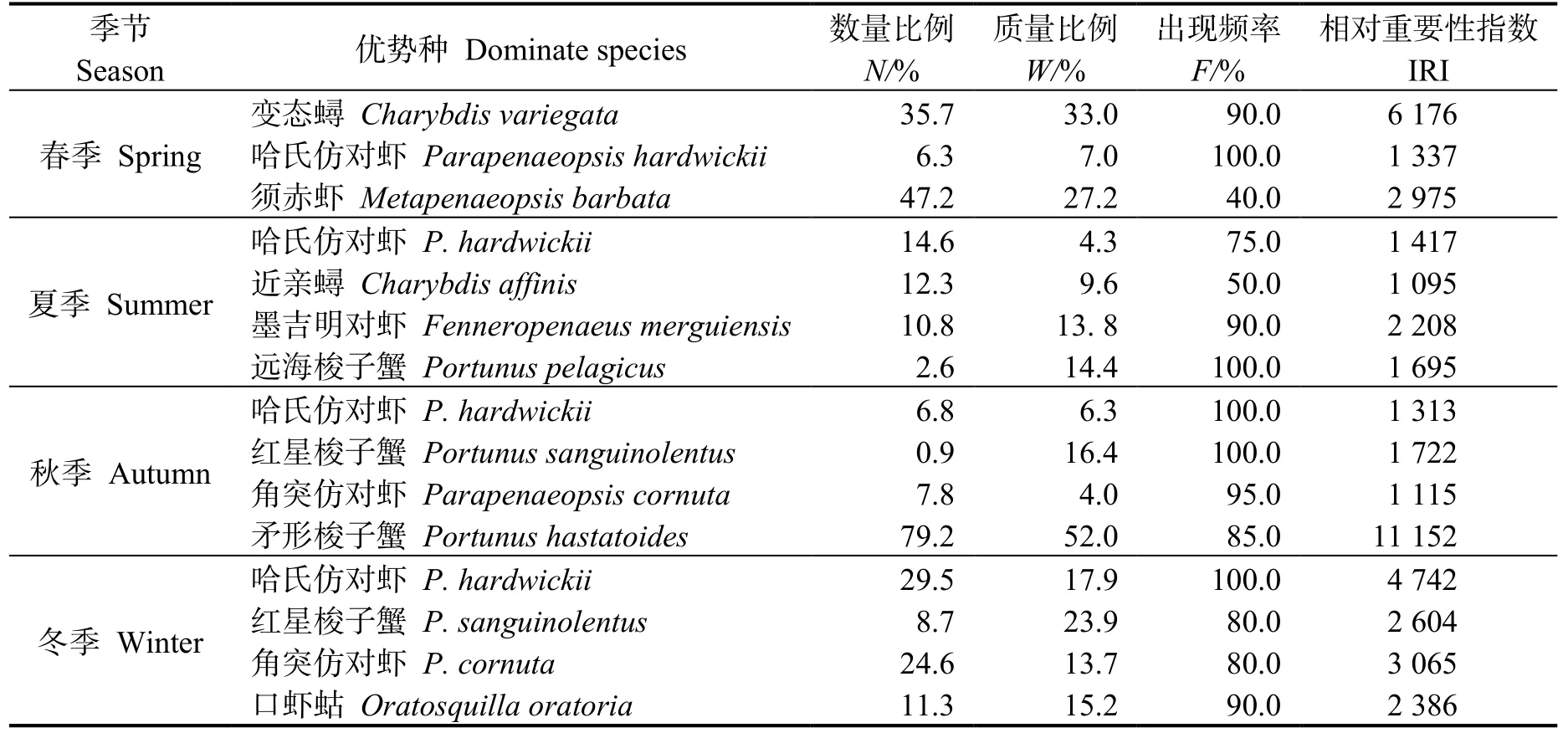
表2 各不同季節優勢種的相對重要性指數(IRI)Table 2 IRI index of dominant species in different season in the east coastal of Leizhou Peninsula

表3 生物多樣性指數的季節變化Table 3 Seasonal change of the diversity index in the east coastal of Leizhou Peninsula
四個季節甲殼動物均可劃分為2個群落(圖2),春季群落Ⅰ主要分布在10 m 等深線附近,以哈氏仿對蝦、遠海梭子蟹(Portunus pelagicus)、紅星梭子蟹等為主;群落Ⅱ主要分布在20 m 等深線附近,以須赤蝦、口蝦蛄、伍氏平蝦蛄(Erugosquilla woodmasoni)等為主;夏季群落Ⅰ位于該海域的西南一側,靠近雷州半島,以近親蟳、遠海梭子蟹、墨吉明對蝦等為主;群落Ⅱ位于東側及北部,以猛蝦蛄(Harpiosquilla harpax)、遠海梭子蟹、墨吉明對蝦等為主;秋季群落Ⅰ位于西南一角的淺灘范圍,以紅星梭子蟹(P.sanguinolentus)、遠海梭子蟹、哈氏仿對蝦等為主;群落Ⅱ以矛形梭子蟹、紅星梭子蟹、哈氏仿對蝦等為主;冬季群落Ⅰ主要位于南部,以紅星梭子蟹、角突仿對蝦、哈氏仿對蝦等為主;群落Ⅱ主要位于20 m 等深線附近,以口蝦蛄、伍氏平蝦蛄、哈氏仿對蝦等為主。
2.5 群落相似性與相異性
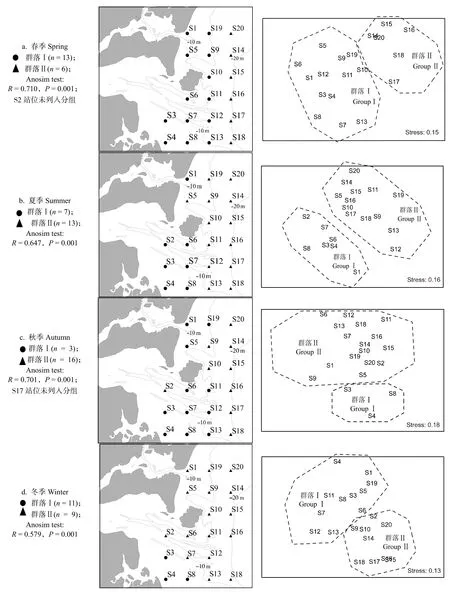
圖2 不同季節甲殼動物的聚類分析和MDS 排序Fig.2 Cluster analysis and MDS sequence diagram of crustaceans in different seasons in the east coastal of Leizhou Peninsula
表4 表明,春季群落I、II 的組內相似性分別為44.5%、54.8%,群落Ⅰ主要特征種為哈氏仿對蝦等,群落Ⅱ主要特征種為須赤蝦等。夏季群落Ⅰ、Ⅱ的組內相似性分別為54.4%、53.8%,群落Ⅰ主要特征種為近親蟳等,群落Ⅱ主要特征種為遠海梭子蟹等。秋季群落Ⅰ、Ⅱ的組內相似性分別為62.7%、59.8%,群落Ⅰ主要特征種為哈氏仿對蝦等,群落Ⅱ主要特征種為紅星梭子蟹等。冬季群落Ⅰ、Ⅱ的組內相似性分別為47.3%、53.8%,群落Ⅰ主要特征種為哈氏仿對蝦等,群落Ⅱ主要特征種為哈氏仿對蝦等,貢獻率前5 位的物種見表4。
春、夏、秋、冬四季群落Ⅰ與群落Ⅱ的相異性(表5)分別為70.6%、58.1%、50.2%和63.0%。春季群落間的主要分歧種為須赤蝦等,夏季為近親蟳等,秋季為猛蝦蛄等,冬季為伍氏平蝦蛄等。貢獻率前5 位的物種見表5。

表4 不同季節群落平均相似性及其貢獻率前5 位的種類Table 4 Top five representative species and their contributions to the average within-group similarity in different seasons in the east coastal of Leizhou Peninsula %
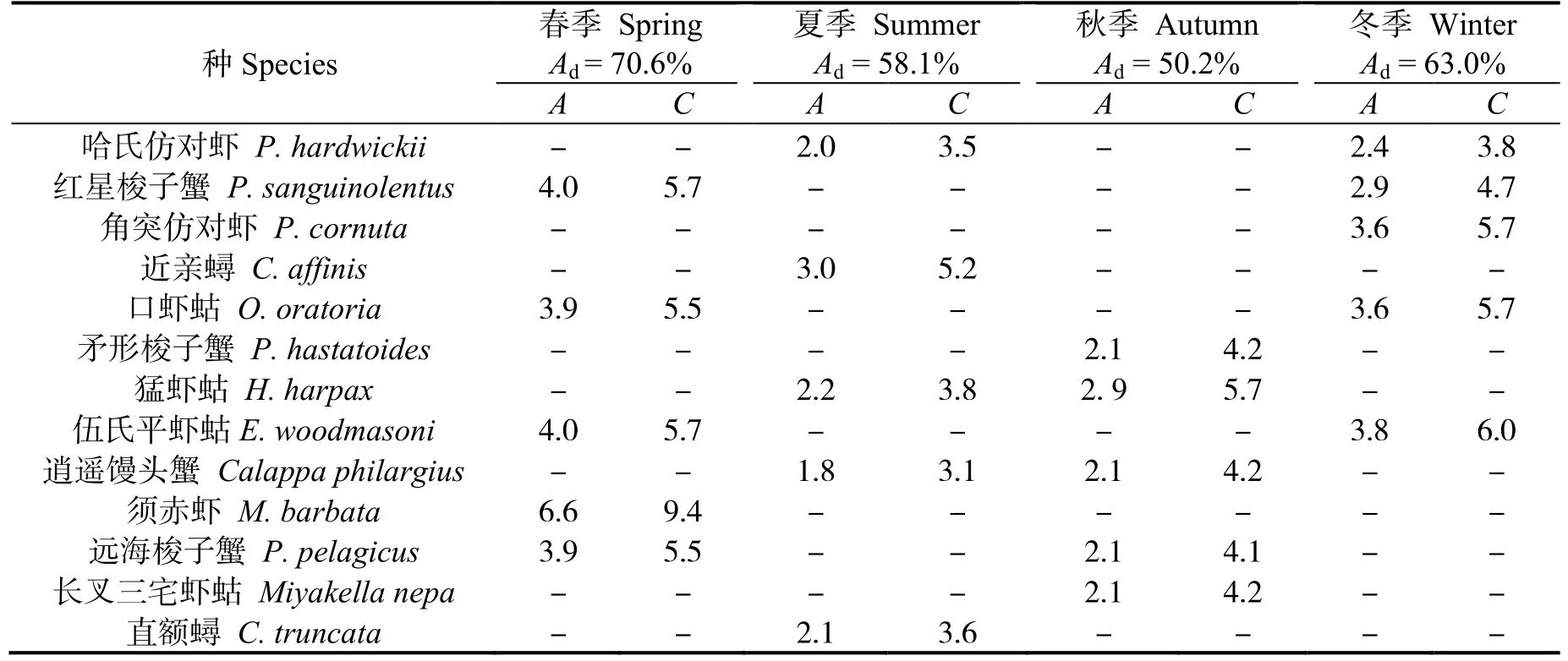
表5 各季節群落間甲殼動物平均相異性及其貢獻率前5 的種類Table 5 Top five representative species and their contributions to the average between-group dissimilarity in different seasons in the east coastal of Leizhou Peninsula %
2.6 群落結構與環境因子的關系
DCA 分析表明,梯度長度的最大值均小于3,因此選用RDA 排序方法。當各季節甲殼動物群落-環境多元回歸線性關系的R2經Ezekiel 公式[21]校正后的值無限接近于零或為負數時,環境因子對甲殼動物群落的解釋能力不能優于初始時隨機生成正態分布的解釋變量[22],因此采用初始RDA 分析結果(表6)。
甲殼動物不同季節物種-環境相關性在RDA1軸(特征值e=1.7223~4.0220)、RDA2 軸(特征值e=0.8675~ 1.9030)分別為44.3%~ 51.3%、15.8%~ 26.2%(圖3)。
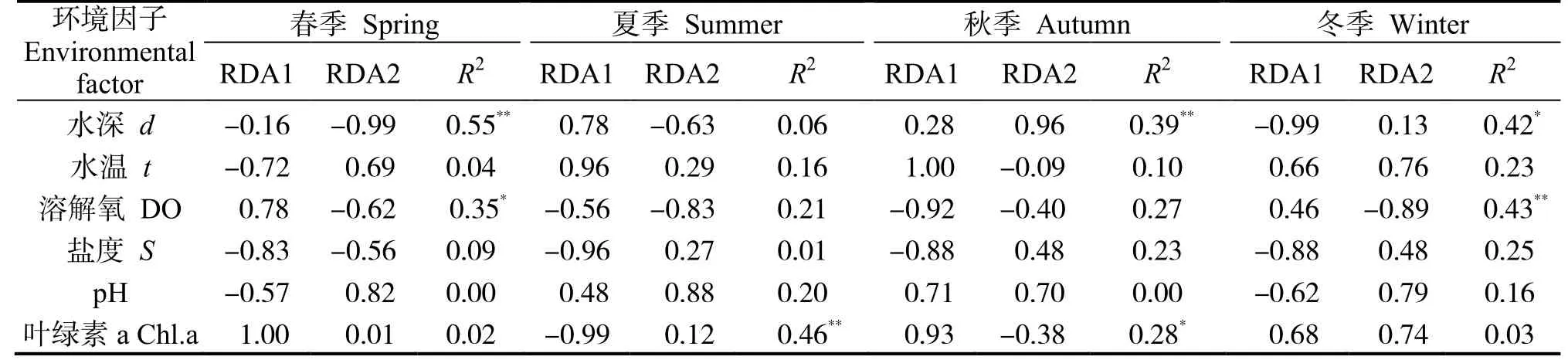
表6 環境因子的相關顯著性檢驗Table 6 Significance test of environmental factors
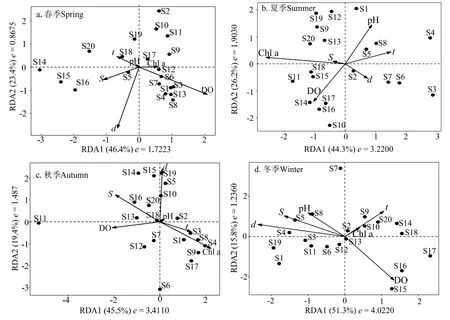
圖3 群落分布與環境因子的RDA 排序Fig.3 RDA triplots of crustaceans community and environmental factors
春季各環境因子對各站位甲殼動物群落結構(種類組成與密度)的貢獻率為32.8%,其中水深與溶解氧是主要影響因子。水深對群落Ⅱ中S14、S15 和S16 站位的甲殼動物分布有極顯著影響,呈正相關,對群落Ⅰ靠近雷州灣及湛江港出港口位置甲殼動物分布的影響呈正相關。溶解氧對群落Ⅰ南側站位的甲殼動物分布影響較大,呈正相關,對群落Ⅱ除S17 外其他站位的影響呈正相關。
夏季各環境因子對甲殼動物群落結構的貢獻率為31.6%,其中,葉綠素a 是主要影響因子。葉綠素a 對群落Ⅱ甲殼動物分布有極顯著的影響,呈正相關,對群落Ⅰ東側站位的影響呈正相關。
秋季各環境因子對甲殼動物群落結構的貢獻率為30.8%,其中,水深與葉綠素a 是主要影響因子。葉綠素a 對群落Ⅰ甲殼動物分布有極顯著的影響,呈正相關,對群落Ⅱ S1、S9、S2、S6、S7 和S12 站位的影響呈正相關。水深對群落Ⅰ甲殼動物分布的影響呈負相關,對群落Ⅱ S2、S5、S10、S13和東側站位的影響呈正相關。
冬季各環境因子對甲殼動物群落結構的貢獻率為39.5%,其中,溶解氧與水深是主要影響因子。溶解氧對群落Ⅰ S1、S6、S12、S13 站位甲殼動物分布的影響呈正相關,對群落Ⅱ S14、S15、S16、S17 和S18 站位有極顯著的影響,呈正相關。水深對群落Ⅱ的影響呈正相關,對群落Ⅰ除S13 外其他站位的影響呈正相關。
3 討論
3.1 甲殼動物群落結構特征
3.1.1 種類組成 本研究發現甲殼動物的種類數(98 種)與鄰近緯度地區的珠江口附近海域[23](54種)、大亞灣口海域[24](52 種)和柘林灣海域[25](53種)以及中緯度地區的樂清灣海域[26](43 種)、蒼南近海海域[27](45 種)和舟山海域[4](42 種)相比,更具優勢。一般認為,低緯度地區的生物多樣性水平高于高緯度,且捕撈強度是重要因素。研究海域處于北回歸線以南的低緯度地區,受熱帶和亞熱帶季風氣候影響,溫度、鹽度穩定,日照時間長,餌料生物資源充足,因此種類數亦較豐富[9-12]。此外,亦與研究海域的面積、海底地形和航次等因素有關。
雷州半島東部近岸海域全年優勢種有10 種,包括生態類型為暖水廣鹽性的變態蟳、近親蟳、紅星梭子蟹、墨吉明對蝦、遠海梭子蟹和矛形梭子蟹,廣溫廣鹽性的哈氏仿對蝦、口蝦蛄、須赤蝦和角突仿對蝦。優勢種季節更替方面,春-夏季變化較顯著,更替率達50.6%,其他相鄰季節變化也較明顯。可能是春-夏季交替期間,溫度回升,甲殼動物活動頻繁,且正值多數甲殼動物的產卵高峰期,如哈氏仿對蝦主要汛期為2―5 月和8―11 月,矛形梭子蟹5―6 月常見抱卵。其中,哈氏仿對蝦在各個季節均為優勢物種,可能與其生命周期短、繁殖能力強和生長迅速等特性有關[28]。
3.1.2 生物多樣性 多樣性指數 (H′) 常作為評價水域生態環境健康的指標之一,也用于評價生物群落受自然因素和人為因素共同影響的程度(0<H′<1 為重度影響;1<H′<3 為中度影響)[29]。本研究中,甲殼動物多樣性指數 (H′) 均值為2.37,說明其群落分布受到中等程度的自然、人為因素影響;豐富度指數 (D) 季節均值為4.77,比柘林灣[25](D=2.45)、樂清灣[26](D=1.64)和舟山海域[4](D=1.37)等物種豐富程度高,可能與該海域的自然環境優越、生物群落穩定性高和人類選擇性捕撈魚類過度等因素有關。四季中以夏季的生物多樣性指數均為最高,而秋季的生物多樣性水平卻最低。夏季正值休漁季節,海洋生物受人類干擾程度最小,生物多樣性水平較高;秋季正值伏季休漁結束,開漁期人們在巨大利益驅動下對海洋生物資源影響最大,致使該季節生物多樣性水平較差。這充分說明伏季休漁制度對甲殼動物資源具有重要養護作用,雷州半島東部近岸海域甲殼動物資源及生態環境的跟蹤監測與休漁結束后的漁業管理政策需要加強,以促進休漁效果的累積和資源的合理開發。
3.1.3 群落劃分 雷州半島東部近岸海域甲殼動物群落結構四季變化明顯,四個季節均可劃分為兩個群落,MDS 二維標序圖、Anosim test 和Simper test結果均表明群落劃分的合理性。四季中,除春季群落組間相異性遠高于組內相似性外,其余三季節差距均較小。春季群落Ⅱ以須赤蝦等為主,主要特征種亦是須赤蝦,且群落組內分歧種為須赤蝦,表明須赤蝦對春季群落劃分直接相關。
3.2 甲殼動物群落結構與環境因子的關系
RDA 將對應分析與多元回歸分析相結合,用以顯示物種群落與環境因素之間關系[29]。本研究RDA 排序結果表明,水深、溶解氧和葉綠素a 是不同季節影響該海域甲殼動物群落分布的主要環境因素,且以水深的影響較大。
水深與溶解氧是春季和冬季甲殼動物群落的主要影響因子。冬季溫度相對較低,海水溶解氧隨溫度的降低而升高,甲殼動物開始從沿岸淺灘往深水區遷徙;春季氣候回暖,海水溶解氧開始降低,且實測數據顯示,冬、春兩季溶解氧含量相差較大,促使甲殼動物往沿岸淺水區域洄游。甲殼動物營底棲生活,隨所處水深的環境不同,海洋的理化性質(溫度、鹽度等)亦會產生較大的變化[9],而溶解氧限制其生理耐受性[31],影響了甲殼動物棲息地的選擇與生活方式,導致不同地域甲殼動物的生物量和群落分布有較大差異。受限于實測數據,本研究僅有表層水溫、鹽度數據,可能造成結果有一定誤差,但考慮到同一斷面的綜合影響,亦有相近研究將表層溫、鹽度數據納入影響范圍,故未將其剔除,后續調查應增加底層環境因子的監測。
葉綠素a 是影響夏、秋季甲殼動物群落分布的主要環境因素。雷州半島東部近岸海域三面環陸,海底地形復雜、地質穩定,這為底棲生物提供良好的孕育場[32],研究海域日照時間充足,夏、秋兩季日照時長尤其明顯,且受到近年來氣候的變化和環境污染(工、農業發展、海水養殖)等因素影響,初級生產力資源豐富甚至達到水體富營養化水平,進而驅動著次級生產力的生物量變化[9,11,33],最終對甲殼動物的資源變動、群落分布產生影響。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
