尾加壓素Ⅱ在高糖誘導心肌細胞氧化應激、內質網應激中的作用及機制研究
2021-02-04 01:18:18支杏敏張捷盧東戴紅艷
中國循環雜志 2021年1期
支杏敏,張捷,盧東,戴紅艷
尾加壓素Ⅱ是一種生長抑素樣神經環肽,最早發現于硬骨魚尾部下垂體中,是目前已知最有效的縮血管活性物質[1]。研究發現,尾加壓素Ⅱ高表達于糖尿病性心肌病(DCM)的心肌組織中[2],提示其可能在DCM發病過程中起重要作用,但具體機制尚不清楚。尾加壓素Ⅱ在多種細胞中表現出誘導氧化應激、活性氧產生的作用[3],如尾加壓素Ⅱ可以通過誘導心臟成纖維細胞、肺動脈平滑肌細胞等產生活性氧,發揮促增殖作用。活性氧已被證實可以引發內質網應激上游信號分子的產生,從而參與調節并啟動細胞凋亡過程[4],而心肌細胞凋亡是DCM發生發展的重要病理生理機制[5]。因此,尾加壓素Ⅱ/氧化應激/內質網應激通路很可能是DCM發病過程中的重要機制之一。本研究為體外實驗,觀察高糖刺激對心肌細胞中尾加壓素Ⅱ及其受體的表達變化,并通過尾加壓素Ⅱ受體拮抗劑Urantide阻斷尾加壓素Ⅱ系統,觀察心肌細胞中氧化應激/內質網應激通道及細胞凋亡相關信號分子表達的改變。
1 材料與方法
1.1 主要試劑和材料
新生1~2 d清潔級Wistar大鼠乳鼠,由山東魯抗醫藥公司提供;羊抗尾加壓素Ⅱ多克隆抗體、羊抗尾加壓素Ⅱ受體多克隆抗體購自美國Sigma公司;兔抗大鼠甘油醛-3-磷酸甘油醛脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)單克隆抗體、兔抗大鼠葡萄糖調節蛋白78(glucose-regulated protein,GRP78)多克隆抗體、小鼠抗大鼠活化轉錄因子6(activating transcription factor 6,ATF6)單克隆抗體、兔抗大鼠CCAAT/增強子結合同源蛋白(C/EBPhomologous protein,CHOP)單克隆抗體、兔抗大鼠半胱氨酸天冬氨酸蛋白酶-12(cysteine-containing aspartate-specific proteases,caspase-12)多克隆抗體購自英國Abcam公司;尾加壓素Ⅱ受體拮抗劑Urantide購自美國Peptides公司;GAPDH以及二氯熒光素二乙酸酯(2',7'-dichlorodihy-drofluorescin diacetate,DCFHDA)購自上海碧云天生物技術有限公司;TAKaRa逆轉錄試劑盒購自日本TaKaRa公司;胎牛血清、細胞培養基(dulbecco's modified eagle medium,DMEM)購自美國Hyclone公司。
1.2 心肌細胞培養
在無菌條件下,取乳鼠心臟剪碎,用0.125%胰蛋白酶反復消化至組織全部消化,收集每次消化后的胰蛋白酶,用含20%胎牛血清的低糖DMEM培養基終止消化。將收集的細胞懸液用200目濾網過濾后離心、重懸,通過差速貼壁方法獲取心肌細胞,接種到35 cm3培養瓶或六孔板上,于37℃,5%二氧化碳(CO2)孵箱培養36 h后進行實驗。
1.3 實驗設計
用低糖DMEM培養基培養心肌細胞36 h后,將培養的心肌細胞隨機分為4組:低糖組(5 mmol/L葡萄糖)、高糖組(25 mmol/L葡萄糖)[6]、高滲組(5 mmol/L葡萄糖+20 mmol/L甘露糖)、高糖+Urantide組,其中高糖+Urantide組用Urantide(10-5mol/L)預刺激45 min,每組6孔,培養24 h后收集心肌細胞。
1.4 細胞內活性氧水平檢測[7]
收集各組心肌細胞,用無血清DMEM培養基清洗3次,加入30 μmol/L DCFH-DA 37℃下孵育20 min,不發光的DCFH-DA可以自由透過細胞膜,細胞內的活性氧可將DCFH-DA氧化成具有熒光的DCFH-DA,其熒光強度與細胞內產生的活性氧的量成正比。再用無血清DMEM培養基洗滌細胞3次,以充分去除未進入細胞內的DCFH-DA。用共聚焦顯微鏡獲得各組細胞圖像。采用image-pro plus 6.0軟件進行熒光密度分析。
1.5 實時熒光定量PCR檢測
按說明用TRIzol提取心肌細胞總RNA,并用紫外分光光度計在260 nm下測量并計算RNA的濃度及純度。根據TAKaRa逆轉錄試劑盒說明,將RNA逆轉錄為互補DNA(cDNA)。采用Promega的SYBR?Green Ⅰ在熒光定量PCR儀上擴增,建立標準曲線,反應步驟:95℃,10 s,35個循環,62℃退火10 s;72℃延伸10 s進行PCR擴增,在每個循環的延伸期收集熒光信號,并通過熔解曲線分析PCR擴增的特異性。所獲數據通過2-ΔΔCt法進行處理,引物序列見表1。

表1 實時熒光定量PCR引物序列
1.6 免疫印跡 (Western blot)檢測
RIPA細胞裂解液與蛋白酶抑制劑Aprotinin按250:1比例消化收集的各組心肌細胞,冰上裂解30 min,劇烈震蕩3次,高速離心機離心后取上清,加入適量蛋白上樣緩沖液,10 min煮沸。用二喹啉甲酸法[8]測定所得蛋白濃度。用10%SDS聚丙烯酰胺凝膠電泳法進行電泳分離,并轉移至聚偏二氟乙烯膜上,室溫下5%脫脂奶粉封閉2 h,然后分別用GRP78多克隆抗體(1:1 000)、ATF6單克隆抗體(1:160)、CHOP單克隆抗體(1:1 000)和caspase-12多克隆抗體(1:1 000)、GAPDH單克隆抗體(1:1 000)4℃搖床孵育過夜,再與相應的辣根過氧化物酶標記的二抗室溫孵育1 h,采用化學發光檢測法(ECL顯色法)處理蛋白條帶。以GAPDH為內參,用Gelpro32軟件對結果進行半定量分析。

圖1 Western blot法檢測高糖刺激對各組細胞內尾加壓素Ⅱ及其受體蛋白表達量的影響(n=3)
1.7 統計學方法
用SPSS19.0軟件進行統計學處理,并用均數±標準差表示。多組數據間的比較采用單因素方差分析,兩組間的數據比較采用成組t檢驗,P<0.05為差異具有統計學意義。
2 結果
2.1 高糖刺激對心肌細胞內尾加壓素Ⅱ及其受體水平的影響(圖1)
Western blot法檢測結果顯示,與低糖組和高滲組相比,高糖組心肌細胞內尾加壓素Ⅱ及其受體蛋白表達量顯著增高(P<0.05)。
2.2 高糖刺激下尾加壓素Ⅱ對心肌細胞內活性氧水平的影響(圖2)
細胞免疫熒光法檢測結果顯示,與低糖組相比,高糖組心肌細胞中活性氧表達水平明顯增高(P<0.0001);與高糖組相比,高糖+Urantide組心肌細胞中活性氧表達水平明顯降低(P<0.0001)。
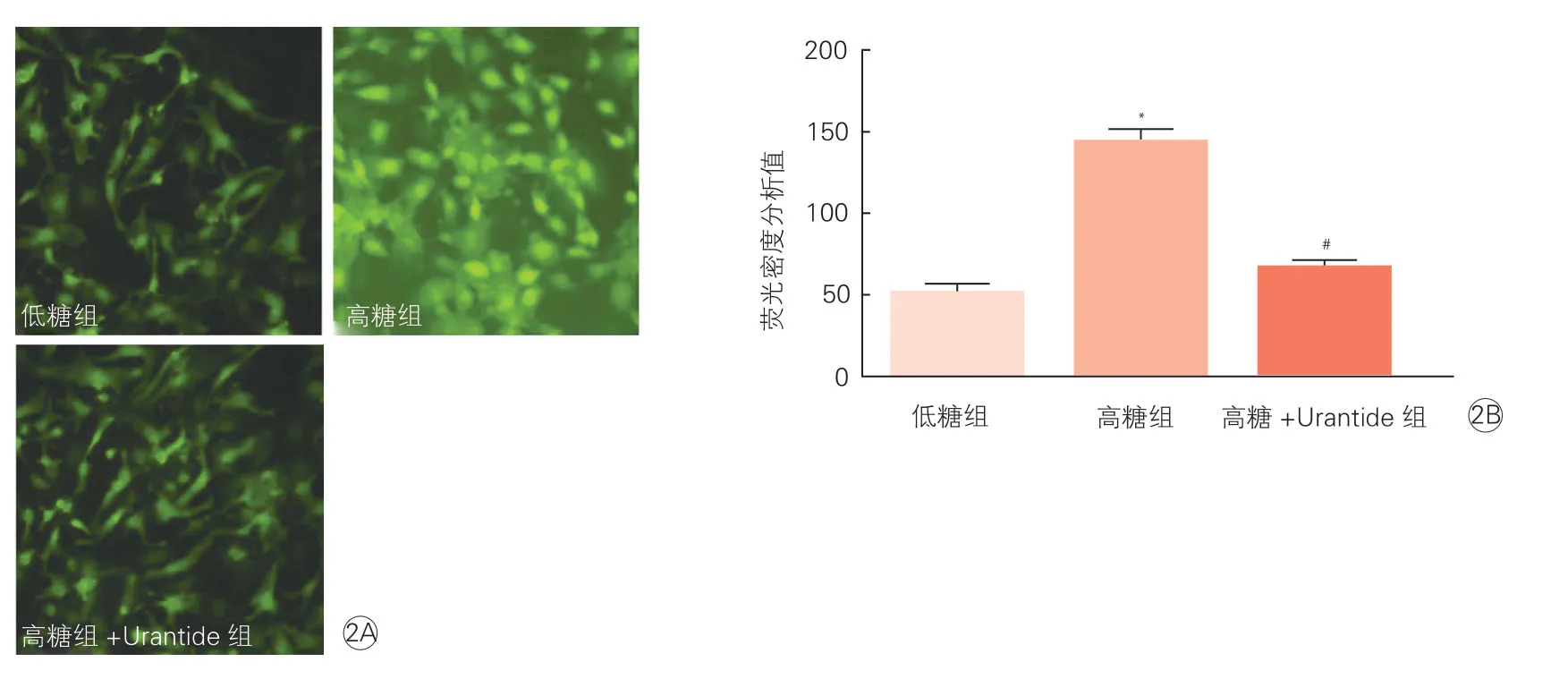
圖2 細胞免疫熒光法分析高糖刺激下尾加壓素Ⅱ對心肌細胞內活性氧水平的影響(n=3)
2.3 高糖刺激及阻斷尾加壓素Ⅱ受體后心肌細胞內質網應激及細胞凋亡相關信號分子的變化(圖3、4)
通過實時熒光定量PCR、Western blot法檢測各組中ATF6、GRP78、CHOP和 caspase-12的表達變化。與低糖組比,高糖組中ATF6、GRP78、CHOP和 caspase-12的mRNA及蛋白的表達量均明顯增高(P<0.05);和高糖組比,高糖+Urantide組中ATF6、GRP78、CHOP和caspase-12的mRNA及蛋白的表達量均明顯降低(P<0.05)。
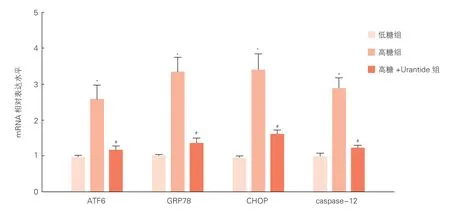
圖3 實時熒光定量PCR檢測高糖刺激下尾加壓素Ⅱ對各組細胞ATF6、GRP78、CHOP和caspase-12的mRNA表達的影響(n=3)

圖4 Western blot法檢測高糖刺激下尾加壓素Ⅱ對各組細胞ATF6、GRP78、CHOP和 caspase-12的蛋白表達的影響(n=3)
3 討論
目前,我國死于糖尿病并發癥的人數比例日益增加,其中DCM已成為加速糖尿病患者死亡的重要原因之一。DCM組織形態學主要表現為心肌細胞凋亡、變性、炎癥反應[9]等,其中心肌細胞凋亡是其最重要的病理改變[5]。研究證實,不論是糖尿病患者[10],還是糖尿病動物模型[5],其心肌細胞凋亡均明顯增加。而采用不同手段降低心肌組織中心肌細胞凋亡后,可預防或減輕DCM患者心功能惡化[11]。因此,心肌細胞凋亡在糖尿病心肌病變過程中發揮了重要作用,但其相關機制尚不完全明了。
尾加壓素Ⅱ是生長抑素類似的環狀血管收縮肽,其縮血管效能可達內皮素-1的10倍[1]。在終末期心力衰竭患者心肌組織及心肌梗死后心力衰竭大鼠心肌組織[2]中,均檢測到尾加壓素Ⅱ及其受體表達增高。DCM時尾加壓素Ⅱ及其受體表達明顯增高[12]。因此,尾加壓素Ⅱ系統的激活很可能參與并調控DCM過程中心肌細胞凋亡。
內質網應激是近年才發現的一種新的凋亡途徑,參與多種疾病及病理狀態下的細胞凋亡。當內質網處于應激狀態時,內質網的分子伴侶如GRP78、葡萄糖調節蛋白94(glucose-regulated protein 94,GRP94)和鈣網蛋白的產生都會相應增加[10],其中GRP78是內質網應激反應的標志蛋白[13],會與錯誤折疊的蛋白或未折疊的蛋白相結合,阻止這些蛋白的合成并使它們在內質網中重新排列折疊或進入降解途徑,以便能夠修復內質網。內質網應激通常可以激活轉錄因子ATF6[14]。ATF6是內質網應激引起細胞凋亡和自噬途徑中的一個重要的調節因子,嚴重或長時間的內質網應激損傷內質網功能時,它能夠啟動由內質網應激所介導的凋亡信號通路,誘導細胞凋亡。內質網應激誘導的下游凋亡信號通路包括[15-17]:CCAAT/增強子結合蛋白同源蛋白或生長停滯及DNA損傷誘導基因153(CHOP/GADDl53)、c-Jun氨基末端激酶(JNK)、天冬氨酸特異性半胱氨酸蛋白酶(caspase)通路。內環境平穩狀態下,CHOP和caspase-12的表達很弱,當穩態失衡時,細胞發生內質網應激,CHOP和caspase-12的表達明顯升高,過度表達的CHOP和caspase-12會促使細胞周期停滯或甚至導致細胞凋亡。
研究證實,內質網應激不但與糖尿病的發病密切相關[18],還參與糖尿病多種并發癥的發生發展,如糖尿病大血管病變、糖尿病腎病及糖尿病視網膜病變。在體外,高糖能誘導血管內皮細胞[19]、大鼠系膜細胞[20]內質網應激的產生;游離脂肪酸通過CHOP通路促進小鼠足細胞內質網應激的激活[21]。內質網應激誘導的心肌細胞凋亡在DCM的發生發展中發揮重要作用。Li等[22]發現,鏈脲佐菌素誘導小鼠心肌組織CHOP、GRP78、GRP94、激活型ATF6及磷酸化α 亞基的真核起始因子2表達均明顯增加,進一步證實了內質網應激在DCM發展過程中的重要作用。體外研究證實,高糖可以通過激活氧化應激誘導心肌細胞內質網應激的產生,進而促進心肌細胞凋亡[23]。
本研究顯示,給予高糖刺激后,心肌細胞中尾加壓素Ⅱ及其受體表達明顯增加,內質網分子伴侶GRP78、轉錄因子ATF6、內質網相關凋亡信號分子CHOP、caspase-12的表達均明顯增加,而尾加壓素Ⅱ受體拮抗劑Urantide可以部分阻止這一過程,提示高糖刺激下尾加壓素Ⅱ可能通過與其特異性受體結合,激活氧化應激、內質網應激通路,進而促進心肌細胞凋亡。
綜上所述,尾加壓素Ⅱ除了具有較強的血管活性外,還可以在高糖刺激下表達增加,進一步激活氧化應激、內質網應激及細胞凋亡通路,從而參與DCM的發病過程。本研究結果豐富了DCM的病理生理學機制,為糖尿病并發癥的防治提供了新思路。
利益沖突:所有作者均聲明不存在利益沖突
